




Chiba Medical J. 99E:17-25, 2023
doi:10.20776/S03035476-99E-2-P17
〔 Chiba Medical Society Young Investigator Award Minireview 〕
Sota Kikuchi1), Atsushi Kaneda2), and Takayuki Hoshii2)
1) School of Medicine, Chiba University, Chiba 260-8670.
2) Department of Molecular Oncology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
(Received September 30, 2022, Accepted January 12, 2023, Published April 10, 2023.)
Histone modification plays an indispensable role in the regulation of transcription. The SET1/MLL family proteins are histone methyltransferases that are conserved from yeast to humans. SETD1A is one of six human SET1/MLL family proteins that primarily catalyzes H3K4 tri-methylation in vitro. SETD1A interacts with multiple subunits to form a complex, the generation of which enhances enzymatic activity and contributes to SETD1Aspecific functions. As H3K4 tri-methylation serves as an active transcription marker, the SETD1A complex is believed to promote the transcription of downstream targets. Over the past decade, considerable work has been conducted on the involvement of SETD1A in human diseases. SETD1A is upregulated in cancers and its elevated expression is correlated with poor prognosis, whereas loss-of-function mutations in SETD1A are associated with neurodevelopmental disorders, such as schizophrenia. A SETD1A knockout mouse model has been characterized by abnormal neurotransmission and neural circuit formation and exhibits schizophrenia-like symptoms. Consequently, SETD1A is assumed to play essential roles in cell proliferation, differentiation, and metabolism. Recently, our group and others have identified the non-catalytic functions of SETD1A, as well as the non-canonical substrates of SETD1A. Importantly, many studies have revealed that SETD1A regulates the DNA damage repair pathway via both canonical and non-canonical routes. However, the underlying molecular mechanisms remain unclear and require further investigation. Here, we review the catalytic and non-catalytic functions of SETD1A in different diseases.
SETD1A, Epigenetics, Cancer, Neurodevelopmental disorders, Non-catalytic
The transcriptional reactions in which RNA is synthesized from DNA are influenced by several factors, among which, the chemical modification of DNA and histone proteins plays significant roles in the regulation of transcriptional processes[1,2]. Histone proteins form a histone octamer consisting of pairs of four core histones (H2A, H2B, H3, and H4) that are wrapped by DNA. Histone tails, which are flexible regions at both ends of the histone fold, are modified with multiple marks such as methylation, acetylation, phosphorylation, and ubiquitination[3,4]. The combination of such chemical modifications contributes to the regulation of transcription and subsequent cellular functions in a complex manner. Among those, methylation, which modifies the lysine and arginine residues of histones H3 and H4, is characterized by three states, namely mono-, di-, and tri-methylation. Whereas monomethylation of histone H3 lysine 4 (H3K4me1) is generally enriched in the enhancer region, H3K4 dimethylation ( H3K4me2) occurs within the gene body, and tri-methylation of H3K4 (H3K4me3) occurs in the vicinity of the promoter region near the transcription start site (TSS) [5,6]. H3K4me3 is assumed to serve as a marker of transcriptional activation and is used to identify the TSS of active genes. In yeast, which are primitive eukaryotes, a single enzyme Set1 catalyzes all H3K4 methylation reactions, whereas in mammals, six Set1 homologous proteins (MLL1-4 and SETD1A/ B) , referred to as SET1/MLL family proteins, have been identified[7,8]. Gene knockout studies have demonstrated functional differentiation among SET1/ MLL family proteins[9-11]. Among Set1 homologs, SETD1A/B, which retain structural similarity, are recognized as methyltransferases for H3K4me3 (Fig. 1) [9]. In recent years, numerous studies based on the use of patient databases and animal models have begun to reveal the diverse roles of SETD1A. In this review, we provide an overview of SETD1A-related diseases and the molecular functions of SETD1A.
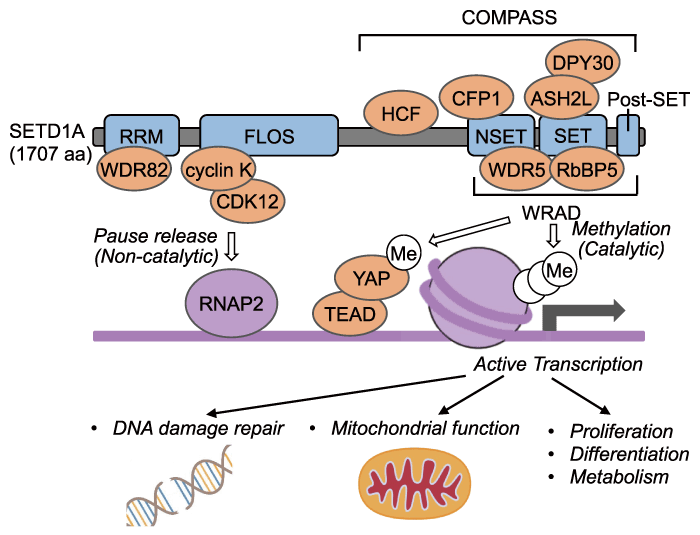
Fig. 1 The scheme of the SETD1A/COMPASS complex. The COMPASS complex catalyzes H3K4me3, resulting in active transcription. It also catalyzes mono-methylation of K342 of YAP, followed by active transcription. FLOS activates transcription via the release of paused RNA polymerase II in a non-catalytic-dependent manner. Abbreviations for domains and proteins are the followings: RRM, RNA recognition motif; WDR82, WD repeat-containing protein 82; FLOS, Functional location on SETD1A; CDK12, Cyclin dependent kinase 12; RNAP2, RNA polymerase II; TEAD, TEA domain transcription factor; SET, Suppressor of variegation 3-9, enhancer of zeste and trithorax; NSET, N-terminal of SET; Post-SET, C-terminal of SET; HCF, Host cell factor C1; CFP1, CXXC finger protein 1; WDR5, WD-40 repeat-containing protein 5; RbBP5, Retinoblastoma-binding protein 5; ASH2L, Ash2 (Absent, Small, Or Homeotic) -like; DPY30, Dumpy-30.
Human SETD1A is a nuclear-localized 1707-amino acid protein that has four evolutionarily conserved domains in common with yeast Set1, namely, the RRM, NSET, SET and post-SET domains. Among these, the RRM (RNA recognition motif) domain is located in the N-terminal region, whereas the NSET, SET and post- SET domains are found in the C-terminal region[12,13]. Methyltransferase activity of SETD1A requires the NSET, SET and post-SET domains. The post-SET domain is a conserved cysteine-rich motif following the SET domain, but the actual role in SETD1A is still unclear. The SET domain is an enzymatically active domain common to all six SET1/MLL family proteins, as well as other histone methyltransferases, including SUV39 and EZH1/2[14]. The NSET and SET domains interact with a conserved complex, called COMPASS (complex of proteins associated with Set1) , and exhibit sufficient enzymatic activity[15-17]. In mammals, four subunits, namely, WDR5, RbBPP5, ASH2L, and DPY30 (referred to as WRAD complex) are common to all SET1/MLL family proteins[16]. SETD1A also interacts with CFP1 (also referred to as CXXC1) , WDR82, and HCF1 to form the COMPASS complex in mammals (Fig. 1) [17]. In addition to the canonical COMPASS complex, SETD1A also forms a complex with the specific subunit BOD1L, which regulates H3K4 methylation followed by DNA damage repair [18]. In contrast, the yeast BOD1L homolog Cps15 is not required for H3K4 methyltransferase activity. Thus, it remains unclear as to whether BOD1L is involved in SETD1A-dependent transcriptional regulation[14]. These protein-protein interactions play an important role in SETD1A-specific downstream regulation, and recent studies have reported that H2B ubiquitination and CFP1 are essential for H3K4me2/3 conversion by SETD1A [19]. The determinants of SETD1A-specific target genes are currently under investigation.
MLL1 is a well-established oncogene involved in pediatric leukemia, whereas MLL3 has been identified as a tumor suppressor in myelodysplastic syndromes (MDS) and acute myeloid leukemia (AML) [20-23]. Although mutations in SETD1A have not been reported or found to be enriched in leukemia and solid tumors, high expression of SETD1A has been reported in several cancers. In the case of MLL-rearranged AML (MLL-r AML) , our group identified SETD1A as an essential factor for cell survival and regulation of the DNA damage repair pathway[24]. Furthermore, in lung, gastric, breast, prostate, and hepatocellular carcinomas, SETD1A is also upregulated and the elevated expression of SETD1A has been demonstrated to be correlated with poor prognosis [25-32].
Several reports indicate SETD1A involvement in cancer development and progression. The Wnt/β-catenin pathway is associated with stem proliferation and cell pluripotency[33,34]. In non-small cell lung cancer, SETD1A activates the Wnt/β-catenin pathway via H3K4me3 activation in the promoter regions of EZH2 and the lncRNA NEAT1, thereby repressing AXIN2, GSK3β, and ICAT, which are negative regulators of the Wnt/β-catenin pathway. Moreover, β-catenin has been found to positively regulate SETD1A via TCF4. Thus, SETD1A and the Wnt/β-catenin pathway is assumed to form a positive feedback loop[35]. Moreover, knockdown of SETD1A has been shown to reduce the expression of SOX2, OCT4, CD133, and other cancer stem cell-associated genes and increases sensitivity to cisplatin, the standard chemotherapeutic agent administered for lung cancer [35]. In addition to activation of the Wnt/β-catenin pathway, SETD1A promotes epithelial-mesenchymal transition (EMT) , which is associated with tumor migration, invasion, and metastasis. Suppression of SETD1A increases the epithelial cell marker CDH1 and reduces the certain mesenchymal cell markers (CDH2, ZEB1, phosphorylated SMAD2, and SNAI1). Xenograft-based studies have revealed that suppression of SETD1A results in the suppression of tumor metastasis in nude mice[25], and SETD1A has similarly been observed to enhance EMT in gastric cancer [36]and in estrogen receptor α (ERα) -positive breast cancer[28].
In addition, SETD1A assumed to play a role in cancer metabolism. In the case of gastric cancer, SETD1A mediates an enhancement of the glycolytic pathway, in which SETD1A interacts with HIF1α to transcriptionally activate the promoter regions of HK2 and PFK, which are involved in the glycolytic pathway via H3K4me3 activation. Moreover, the suppression of SETD1A results in a decreased glucose uptake and lactate production in gastric cancer cells [27].
SETD1A also contributes to the development of drug resistance in cancers. Tamoxifen resistance has been demonstrated to be correlated with the expression of SOX2, which is highly expressed in breast cancer cells. SETD1A enhances H3K4me3 in the promoter region of SOX2. Furthermore, SETD1A interacts with SOX2 to activate SOX2 target genes in breast cancer cells. Thus, SETD1A-SOX2 axis is assumed to be involved in the tamoxifen resistance in breast cancer [37]. FOXM1 has been reported to be associated with resistance to docetaxel in castration-resistant prostate cancer[38]. SETD1A is recruited to the promoter region of FOXM1 by E2F1, where it promotes transcriptional activation via H3K4me3[29]. A high expression of SETD1A promotes sorafenib resistance in hepatocellular carcinoma via the activation of YAP, a core component of the Hippo signaling pathway, an established tumor suppressor pathway[30]. Collectively, these findings indicate that elevated SETD1A expression is commonly associated with drug resistance in multiple cancers.
In recent years, whole exome sequences have revealed an association between SETD1A and neurodevelopmental disorders, such as autism spectrum disorder and an obsessive-compulsive disorder [39-43]. Heterozygous loss-of-function (LoF) mutations in SETD1A are assumed to contribute significantly to the development of schizophrenia[42], and a Setd1a+/- mouse model demonstrated working memory deficits and impaired learning and social skills[44,45]. These findings are consistent with the symptoms of schizophrenia in patients with SETD1A LoF mutations, and it is accordingly anticipated that studies based on the Setd1a+/- mouse model will contribute to elucidating the etiology of schizophrenia. Interestingly, overexpression of SETD1A or pharmacological antagonism of LSD1, a demethylase, has been demonstrated to partially rescue the abnormalities in Setd1a+/- mice[44]. It may indicate that SETD1A and its associated cellular functions could serve as potential targets for schizophrenia treatment or other neurodevelopmental disorders.
SETD1A LoF has been demonstrated to show functional abnormality in neuron. With the exception of cortical layer L1 (the outermost layer) , SETD1Apositive cells are distributed throughout the cerebral cortex. In Setd1a+/- mice, attenuation of excitatory synaptic transmission has been detected in cortical L2/3, which is assumed to be associated with a reduction in presynaptic glutamate release and reduced postsynaptic receptor density[45]. In a further study, Setd1a+/- mice were found to be characterized by the downregulated expression of mitochondria-related genes involved in glutamine metabolism[46]. In addition, SETD1A LoF mutation leads to fluctuation of glutamatergic neuron-related genes and an increase in the burst activity of excitatory neuronal networks in human iPSC-derived neurons [47]. These findings thus tend to indicate that SETD1A dysfunction may result in an imbalance between neuronal glutamate supply and demand. Such abnormalities are, however, not limited to synaptic transmission, as revealed by the observation that SETD1A LoF mutation also induces changes in the pattern of nerve axon projections. For example, during cortical development, abnormal axon projections can contribute to disorders in the formation of neural circuits. These changes in neuronal cell functions are considered to be associated with higher brain dysfunction and lead to neuronal circuit failure, resulting in short-term synaptic plasticity[44], which is linked to learning and memory[48]. This potentially accounts for symptoms such as learning disabilities in Setd1a+/- mice and SETD1A LoF mutation patients.
In addition, SETD1A is assumed to be essential in early stage of neuron development. SETD1A is reportedly associated with early-onset neurodevelopmental disorder, such as early onset epilepsy[43], and functional analysis of SETD1A in neural progenitor cells is currently underway. SETD1A contributes to cell proliferation and differentiation in mouse embryonic neural progenitor cells by activating the Wnt/β-catenin pathway[33,49]and interacting with the histone chaperone Nap1l1 to activate RassF10 via H3K4me3[50]. Furthermore, in human neuroblastoma cells, suppression of SETD1A causes the upregulated expression of DCX, which is associated with neural migration, and the downregulated expression of DLX5, which plays roles in the development of cortical interneurons[51]. These findings suggest that SETD1A plays an important role in the differentiation and functions of neuron.
The findings of a number of studies have implicated SETD1A in immune responses. In activated murine macrophages, for example, glucocorticoid receptor (GR) recruits the SETD1A/COMPASS complex in the enhancer regions of genes downstream of GR. The SETD1A/COMPASS complex is required for the anti-inflammatory effects following the GR-mediated transcription. In contrast, no significant correlation has been detected between H3K4 methylation and the GR-mediated transcriptional control of inflammatory response genes[52]. Consequently, these findings appear to indicate involvement of the non-enzymatic functions of SETD1A in the immune system, which are described below.
In addition to inflammation, coagulation is also an innate immune response. Moreover, overactivation of the immune system often leads to clinical problems such as thrombosis [53,54]. In this regard, SETD1A has been established to activate the transcription of THBD, which encodes thrombomodulin that is associated with thromboresistance in vascular endothelial cells via H3K4me3[55]. Thus, SETD1A may broadly contribute to immune system activity and microenvironment in human diseases.
Recently, several non-canonical substrates of SETD1A have been identified. For example, activation of YAP in the Hippo signaling pathway in hepatocellular carcinoma (see Section III) has been established to be regulated by the SETD1A/WRAD complex through the mono-methylation of lysine residue K342 of YAP [56]. Normally, YAP interacts with CRM1 and is subsequently transported out of the nucleus; however, the SETD1A-mediated mono-methylation of K342 inhibits this interaction with CRM1, thereby increasing the YAP nuclear localization and the interaction with TEA domain transcription factor (TEAD). Thus, SETD1A activates YAP/TEAD-dependent transcription and promotes tumorigenesis[56]. SETD1A has also been found to interact with the E3 ubiquitin ligase protein RAD18, which activates the DNA damage repair pathway. However, this interaction occurs independently of the COMPASS components RbBP5 and ASH2L [57]. In addition, as mentioned in Section III, SETD1A is required for the proliferation of MLL-r AML cells in a SET-domain-independent manner, and in AML cells, the FLOS (functional location on SETD1A) domain located in the central region of the protein is required. A functional rescue study using chimeric constructs of SETD1A and SETD1B indicated that the FLOS domain is essential for SETD1A-specific functions, although other known domains are compatible with these paralogs [24]. The FLOS domain has been established to interact with the cyclin K/CDK12 complex, a kinase associated with RNA polymerase II (RNAP2) C-terminal domain, for the transcriptional activation of genes comprising the DNA damage repair pathway, such as Fancd2 (Fig. 1) [24]. In AML cells, SETD1A target genes are upregulated in a cell cycle-dependent manner, and consequently, inhibition of the non-enzymatic functions of SETD1A effectively suppresses the proliferation of AML cells characterized by a high rate of proliferation [24]. In further experiments, we found that SETD1A target genes involved in mitochondrial function in AML cells. Suppression of SETD1A was found to result in a reduction in mitochondrial function, and subsequent abnormal glutamate metabolism[58]. We also revealed that the SETD1A-cyclin K complex promotes the release of promoter-proximally paused RNAP2 into gene bodies, without affecting the methylations of H3K4 in target genes [58]. Taken together, the non-enzymatic functions of SETD1A appear to be associated with DNA damage repair and the maintenance of mitochondrial function in AML cells. These findings are currently being further investigated in other cancers and neurons.
Epigenetic transcriptional regulation via SETD1Amediated histone methylation has been implicated in the pathogenesis of various diseases. As we have summarized herein, however, several functions of SETD1A that occur independently of histone methyltransferase activity have recently been reported. It is still unclear as to whether these roles are commonly required or are cell context-dependent in cancers, as well as in neurodevelopmental disorders. However, given that SET1/MLL family proteins have a wellconserved SET domain structure and interact with common subunits, it would be challenging to target the SET domain itself or its subunits for SETD1A-specific inhibition. On the basis of the findings reported to date, we predict that identifying novel SETD1A subunits or SETD1A functions will provide insights on novel mechanistic properties that do not overlap with those of other MLL family members, and could thus contribute to the development of novel therapeutic tools for the treatment of SETD1A-related diseases.
S. K. wrote the original draft of the manuscript. A. K. reviewed and edited the manuscript. T. H. conceptualized, reviewed, and edited the manuscript. All the authors read and final version of the manuscript.
T. H. was supported by JSPS KAKENHI Grant Numbers JP22K19538, JP22H03099 and JP22H04684. T. H. was also supported by Mochida Memorial Foundation for Medical and Pharmaceutical Research, SGH Foundation for Cancer Research, and the Leading Initiative for Excellent Young Researchers (LEADER). A. K. was supported by IAAR Research Support Program from Chiba University.
A. K. is a member of the Editorial Board of the Chiba Medical Journal. The other authors declare that there is no conflict of interest to disclose.
Not applicable.
Not applicable.
The authors thank all the laboratory members and collaborators who supported the study.
Address correspondence to Dr. Atsushi Kaneda.
Department of Molecular Oncology, Graduate School of Medicine,
Chiba University, 1-8-1, Inohana, Chuou-ku, Chiba
260-8670, Japan.
Phone/Fax: +81-43-226-2039.
E-mail: kaneda@chiba-u.jp
