




Chiba Medical J. 88E:27~34,2012
doi:10.20776/S03035476-88E-3-P27
[Original Paper]
Mayumi Fujitani, Natalie Grinshtein, Kristen M. Smith and David R. Kaplan
Cell Biology Program and James Birrell Laboratories, The Hospital for Sick Children MaRS Centre, Toronto Medical Discovery Tower, 101 College Street, Room 12-314 M5G 1L7.
(Received November 29, 2011, Accepted January 13, 2012)
Neuroblastoma(NB) is a pediatric tumor that develops from primitive neural crest cells that normally form the sympathetic nervous system. Patients with advanced disease have a poor long-term survival and new therapies are desperately needed. JNJ-10198409 was identified as a potential NB therapy in a screen of a kinase-targeted chemical library. Here we examine the effect of JNJ-10198409 on established NB cell lines( SK-N-AS and IMR 32) and primary NB cell proliferation and cell survival in vitro. JNJ-10198409 selectively inhibited NB cell growth in AlamarBlue assays and rapidly induced apoptotic cell death without affecting the growth of primary pediatric neural crest-like stem cells( SKPs). Signaling analysis suggested that JNJ-10198409 may function by inhibiting the activity of Src family kinases. Future experiments will explore the efficacy of combining JNJ-10198409 with current frontline NB chemotherapeutics.
neuroblastoma, drug discovery
Neuroblastoma(NB) is thought to derive from transformed sympathoadrenal progenitor cells of the neural crest. It is the most common pediatric extracranial solid tumor and accounts for approximately 15% of childhood cancer deaths[1]. High-risk NB is treated with an aggressive therapy regimen that can include chemotherapy, radiation, surgery, bone marrow transplantation and immunotherapy. Unfortunately, the long-term survival of patients older than 1 year with advanced disease is less than 30% due to relapse and extensive metastatic disease[1].
In an effort to discover novel drugs for NB, we screened cells from the bone marrow of high risk NB patients that formed spheres in neural stem cell culture conditions with a collection of 143 compounds that target protein kinases[2], and identified JNJ-10198409 as a compound that potently inhibited cell growth. JNJ-10198409 is a selective PDGFRβ kinase inhibitor that also inhibits c-Abl and Src family kinase activity in vitro[3]. It was developed as an anti-angiogenic compound and was subsequently shown to have anti-proliferative activity against a variety of human cancer cell lines[3,4]. Here, we demonstrate that JNJ-10198409 inhibited the in vitro survival of NB cells and not normal neural crestlike pediatric stem cells, and induced apoptosis of cells from NB patients at nanomolar concentrations. JNJ-10198409 may act in these cells through inhibition of Src family kinases.
Chemicals and reagents. JNJ-10198409[6,7-(dimethoxy-2,4-dihydro-indeno[1,2-c]pyrazol-3-yl)-(3-fluro-phenyl)-amine]was provided by the MedicinalChemistry platform at OICR( the Ontario Institute for Cancer Research).
Cell culture. SKPs(FS105) were isolated from neonatal foreskin and cultured as previously described[5,6] following ethics approval by The Hospital for SickChildren Research Ethics Board. SKPs were grown as spheres in 25 or 75 cm2 flasks in a 37°C, 5% CO2 incubator and passaged every 7 days using 1 mg/mL collagenase. Dissociated cells were replated in serumfree proliferation media( PM: 2% B-27 supplement (Invitrogen), 40 ng/mL basic fibroblast growth factor (bFGF, BD Biosciences), 20 ng/mL epidermal growth factor(EGF, BD Biosciences), 100 U/mL penicillin/streptomycin in 3 : 1 DMEM: F12( Invitrogen))containing 50% SKPs conditioned medium. SKPs were used between passages 1-5 unless otherwise indicated. Neuroblastoma cells(NB88R2, NB12) were isolated from patient bone marrow aspirates as previously described[7]. These lines were found to be partially contaminated with EBV-expressing B-cells(Loen Hansford and D.R. Kaplan, submitted). We therefore established an EBV-free MYCN amplified NB line(NB153) from a patient tumor. While NB88R2 and NB12 are a mixture of NB and B-cells, we found that results are comparable among these 3 lines, so we have chosen to report the results of all three lines.
Tumor samples and bone marrow aspirates were obtained from consented patients, as approved by The Hospital for Sick Children Research Ethics Board. Primary NB cells were passaged every 5 to 7 days by manual tituration and dissociated cells were replated in PM. Cells were used between passages 3-12. Established NB cell lines(SK-N-AS IMR 32) were cultured in PM. NIH-3T3 cells were cultured in DMEM with 10% fetal bovine serum.
Assessment of JNJ10198409 effect on NB lines proliferation. NB cell lines(SK-N-AS and IMR 32)were seeded at 3,000 cells per well in 100 μL medium in 96-well microplates and allowed to adhere for 24 hours prior to drug addition. NB153 spheres were dissociated into single cells and seeded at 10,000 cell per well in 100 μL medium.
JNJ10198409 was dissolved in DMSO to obtain a 50 mM stock solution and serially diluted thereafter for a total of 8-point, 3-fold serial dilutions. Drug effect was compared with cells optimally proliferating in 0.05% DMSO alone, whereas wells filled with media served as background. AlamarBlue( 10 μL) was added after 72 hours, and fluorescence intensity was measured after 24 hours on a PHERAstar microplate reader, equipped with a 540 excitation/l590 emission filter.
Sphere-formation assay. Cells were seeded in triplicate in 96-well non-tissue culture-treated plates at 3,000 cells/well(2,000 cells/well for NB88R2 cells) in 50 μL of PM+CM. Compounds were diluted in PM+CM to the indicated concentrations and immediately added to seeded cells in a volume of 50 μL, bringing the final volume to 100 μL. Wells were retreated with compound at the indicated concentrations 3 days postplating. Cultures were fixed with 4% paraformaldehyle (Electron Microscopy Sciences) at day 7 and sphere number determined manually. Percentage of control sphere number was calculated as( mean sphere number for treated wells/mean sphere number of 0.05% DMSO-treated wells) x100. EC50 curves were generated with data from two individual NB cell samples and one SKP sample using GraphPad Prism software(GraphPad Software, Inc.).
AlamarBlue® cell proliferation assay. Cells were seeded and treated with the compounds as above. At day 6, alamarBlue® was added to 10% and the plates were incubated an additional 24 hours. Fluorescence intensity was measured on day 7 with an excitation filter of 535nm and an emission filter of 590 nm. EC50 curves were generated using GraphPad Prism software.
Immunostaining. Three million dissociated NB88R2 cells were plated in 75 cm2 flasks in 30 mL PM. JNJ-10198409 was added to a final concentration of 0.1 μM or 1 μM. After 3 days, spheres were collected, deposited on slides by cytospin and stained with anti-Ki67 antibody(BD Bioscience, 1 : 200) or anti-cleaved caspase 3 antibody(Cell Signaling #9661, 1 : 200). Total Ki67-positive cells, Ki67-negative cells with no cleaved caspase 3 or condensed nuclei and Ki67-negative, cleaved caspase 3-positive cells with condensed or fragmented nuclei were quantified. Cells from 6 random fields were quantified using a 40X objective.
Western blotting. NB88R2 cells were dissociated and plated in 75 cm2 flasks at 3x106 cells in 30 mL PM with 0.1 μM JNJ-10198409, 1 μM JNJ-10198409 or 0.01% DMSO for 48 hours. Cells were collected, washed in cold phosphate-buffered saline, and lysates were prepared in NP-40 lysis buffer(20 mM Tris pH 8.0, 137.5 mM NaCl, 1% NP-40, 20 mM NaF, 0.5 mM Na3VO4, 1 mM PMSF, 20 μg/mL aprotinin, 10 μg/mL leupeptin). Equal amounts of protein(25-50 μg) were resolved on 10% polyacrylamide gels and subjected to immunoblotting with the following antibodies:anti-phospho-PLCγ1(#2821, 1 : 1000), anti-PLCγ1 (Abcam ab16953, 1 : 4000), anti-phospho-Shc(#2431, 1 ; 1000), anti-phospho-Akt( #9271, 1 : 1000), antiphospho-Src(#2101, 1 : 1000), anti-phospho-Erk (#9101, 1 : 2000), anti-cleaved human PARP(Poly ADP ribose polymerase)(#9541, 1 : 1000) and antip38MAPK(#9212, 1 : 1000). Unless otherwise indicated, all antibodies were from Cell Signaling Technology. HRP-conjugated goat anti-mouse IgG and goat anti-rabbit IgG secondary antibodies were used.
PDGF stimulation. NIH-3T3 cells were plated in 10 cm plastic dish. After the cells had reached 80-90% confluency, the cell media was replaced with DMEM plus 0.5% fetal bovine serum for 24 hours. Serumstarved cells were pretreated with DMSO or 5 μM JNJ-10198409 for 30 minutes followed by a 10 minute treatment with 100 ng/mL PDGF or vehicle(4mM HCl). Cells were washed twice with cold PBS, lysed in 300 μL NP-40 lysis buffer and immunoblotted as described above. One million dissociated NB88R2 cells were plated in 25 cm2 flasks in DMEM: F12 with 2% B27. After 3 hours, cells were pretreated with DMSO or 5 μM JNJ-10198409 for 30 minutes followed by a 10 minute treatment with 100 ng/mL PDGF, EGF and bFGF. NB88R2 cells were washed, lysed and processed as described for NIH/3T3 cells.
Immunoprecipitation. NB88R2 cells were dissociated and plated in 75 cm2 flasks at 3x106 cells in 30 mL PM with 0.1 μM JNJ-10198409 or 0.01% DMSO for 48 hours. Treated cells were lysed as above and 250-500 μg of protein lysate were incubated with anti-phosphotyrosine (4G10, Upstate Biotechnology) or mouse IgG(Santa Cruz Biotechnology). Immune complexes were collected with protein A-Sepharose, washed in lysis buffer, subjected to SDS-PAGE and immunoblotted with anti-c-Src(Cell Signaling #2109).
JNJ-10198409 inhibits neuroblastoma cell growth in vitro. Previously, we developed a cell-based high-throughput assay to identify novel therapeutic agents that are selectively cytotoxic or cytostatic against NB cells[8]and utilized this assay to screen a collection of 143 kinase inhibitors[2]. From this screen, we identified JNJ-10198409(Fig. 1A), a PDGF receptor β tyrosine kinase inhibitor as cytotoxic against cells from multiple NB patients. To confirm the effect of JNJ-10198409 on NB cell lines and primary NB cell(NB153) proliferation, we examined cell growth using an alamarBlue assay and obtained EC50 values for each cell line(Fig. 1B)(Figure 1B; SK-N-AS: EC50=20 nM IMR32: EC50=10 nM; NB153: EC50=92 nM). We then assessed in vitro sphere formation capacity as a combined measurement of cell survival and proliferation activity[8]. Sphere-forming cells obtained from the bone marrow of two high-risk NB patients(NB12, NB88R2) were dissociated, plated in triplicate in 96- well plates and treated with serial dilutions of JNJ-10198409. Cells were re-treated with the compound on day 3 and sphere number was determined at day 7.In addition, we treated normal human pediatric skin-derived precursor cells(SKPs). SKPs are a neural crest-like stem cell population that can be cultured from pediatric dermis and share many properties with primary NB sphere-forming cells including self-renewal as spheres in vitro and expression of neural crest markers as well as differentiation into neural crest lineages. However SKPs are non-tumorigenic and thus provide a normal non-transformed stem cell population to evaluate whether JNJ-10198409 is selectively toxic against NB cells. Treatment with JNJ-10198409 had no effect on SKPs sphere formation in vitro at any dose tested (Fig. 1B) but inhibited sphere formation of both NB patient cell isolates in a dose-dependent manner. The EC50 for sphere formation was 283 nM for NB12 cells (95% CI(Confidence Interval) 29.46 to 2729 nM) and 68 nM for NB88R2 cells(95% CI 35.51 to 126.7 nM). These results demonstrate that JNJ-10198409 selectively targets the survival and/or proliferation of NB cell lines and cells from NB patients in vitro.
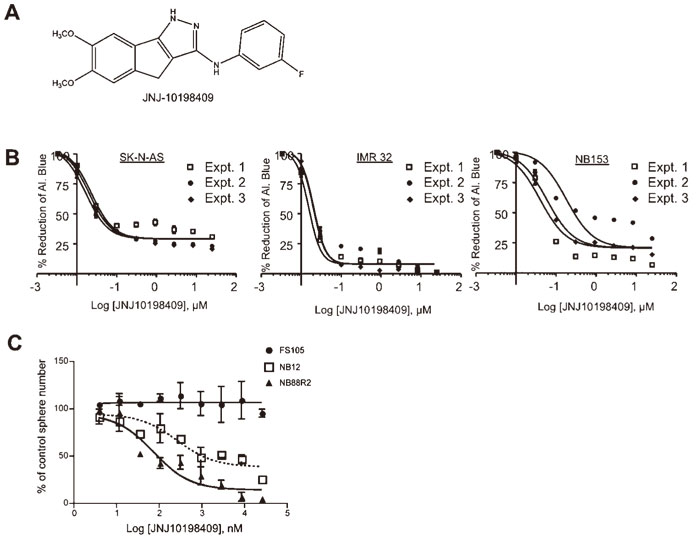
Fig. 1
JNJ-10198409 inhibits NB cell growth at nanomolar concentrations.
A, Chemical structure of JNJ-10198409.
B, JNJ-10198409 inhibits proliferation of NB cell lines SK-N-AS and IMR32 and a primary NB sphere-forming line NB153. EC50 curves are shown demonstrating the effect of JNJ-10198409 on the proliferation of NB cell lines, using AlamarBlue as a read-out.(SK-NAS:EC50=20 nM, IMR32: EC50=10 nM, NB153: EC50=92 nM)
C, EC50 curves are shown demonstrating the effect of JNJ-10198409 on the self-renewal and survival of cells from the bone marrow of high risk NB patients as measured in a sphere formation assay. NB cells(NB12 and NB88R2) had a dose-dependent decrease in sphere formation with EC50 values in the nanomolar range(283 nM for NB12 and 68 nM for NB88R2) while normal pediatric neural crest-like stem cells(SKPs, line FS105) were unaffected. Points, mean of 2 independent experiments; Bars, SD.
JNJ-10198409 treatment induces rapid cell death via apoptosis. JNJ-10198409 has been reported to have anti-proliferative effects on various cancer cell lines in vitro. To investigate the mechanism of action of JNJ-10198409 in cells from NB patients, NB88R2 cells were treated with 0.1 μM, 1 μM JNJ-10198409 or DMSO for 72 hours. Treated cells were immunostained for markers of proliferation(Ki67) and apoptosis(cleaved caspase 3, nuclear DNA condensation and fragmentation). Ki67-positive cells and Ki67-negative cells without cleaved caspase 3 staining or condensed nuclei were scored as live cells while apoptotic cells were identified as Ki67-negative, cleaved caspase 3-positive cells or Ki67-negative cells with condensed or fragmented nuclei(Fig. 2A). Under the control conditions, NB88R2 cells grew robustly as spheres and the majority of cells were Ki67-positive or lacked markers of apoptotic cell death(Fig. 2B). Low concentrations of JNJ-10198409 induced rapid apoptosis in NB88R2 cells with cleaved caspase 3 and condensed or fragmented nuclei observed in 66.2+6.8% of cells. JNJ-10198409 treatment induced apoptosis in a dose-dependent manner with over 83.7+9.8% of cells treated with 1 μM JNJ-10198409 exhibiting markers of apoptosis by 72 hours. Additionally, treatment with 0.1 μM JNJ-10198409 induced an increase in PARP cleavage after 48 hours, supporting apoptosis induction via activation of the caspase cascade(Fig. 2C).
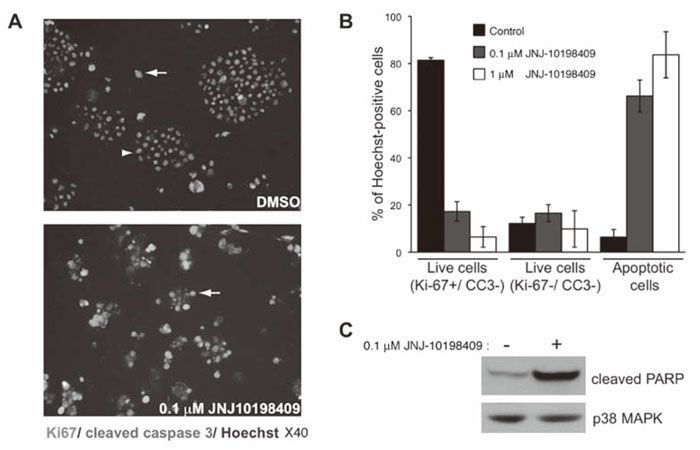
Fig. 2
JNJ-10198409 induces apoptotic cell death of cells from NB patient’s bone marrow.
A, NB88R2 cells were treated with DMSO, 0.1 μM JNJ-10198409 or 1 μM JNJ-10198409 for 72 hours. Treated cells were deposited on slides by cytospin and subjected to immunostaining for Ki67(red) and cleaved caspase 3(green) with Hoechst 33342-staining of nuclei(blue). A representative field is shown for the indicated treatments.
B, Quantification of live and apoptotic cells from 6 random fields for each treatment condition as described in Methods.
C, NB88R2 cells were treated with 0.1 μM JNJ-10198409 or DMSO for 48 hours. Immunoblots of JNJ-10198409-treated cell lysates demonstrated increased PARP cleavage. Total protein loading was visualized by p38MAPK immunuoblotting.
JNJ-10198409 does not act through PDGFRβ inhibition in cells from NB patients but inhibits Src phosphorylation. JNJ-10198409 was developed as a PDGFRβ selective antagonist and both primary NB tumors and some established NB cell lines have been shown to express PDGFRβ[2]. Therefore we asked whether JNJ-10198409 treatment altered PDGFdependent signaling in cells from NB patients. We first investigated whether PDGF-dependent signaling pathways were active in cells from NB patients(Fig. 3A). Stimulation of NIH-3T3 cells with PDGF resulted in rapid phosphorylation of the PDGFR downstream targets, PLCγ1, Shc, Akt and Erk1/2(Fig. 3B). Phosphorylation of the downstream targets was blocked by a 30 minute pre-incubation with 5 μMJNJ-10198409. In contrast, no significant increase in
phosphorylation of PLCγ1, Shc, Akt or Erk1/2 was observed following stimulation of cells from NB patients with PDGF(Fig. 3B). These results suggested that PDGFR was not the target of JNJ-10198409 in those cells.
In addition to PDGFRβ and PDGFRα, JNJ-10198409 inhibits the kinase activity in vitro of c-Abl(IC50=22 nM) and Src family members, c-Src, Lck and Fyn(IC50=100 to 378 nM)[3]. It is unlikely that c-Abl inhibition in cells from NB patients is responsible for the observed cytotoxic effect, because additional c-Abl-selective kinase inhibitors(imatinib and nilotinib) present in the kinase-targeted chemical library were not identified as NB cell-cytotoxic hits at 1 μM[2]. Additionally, imatinib that also inhibits c-Kit and PDGFR kinase activities, had no effect on NB cell sphere formation(K.M.S. and D.R.K., unpublished observations).
To determine whether JNJ-10198409 inhibits Src kinase activity in the cells from NB patients, NB88R2 cells were treated with DMSO or 0.1 μM JNJ-10198409 for 48 hours, and cell lysates were subjected to immunoprecipitation with anti-phosphotyrosine followed by and lysates probing with anti-c-Src(Fig. 3C). Decreased c-Src tyrosine phosphorylation was evident, suggesting that JNJ-10198409 targets Src family kinases in the cells from NB patients.
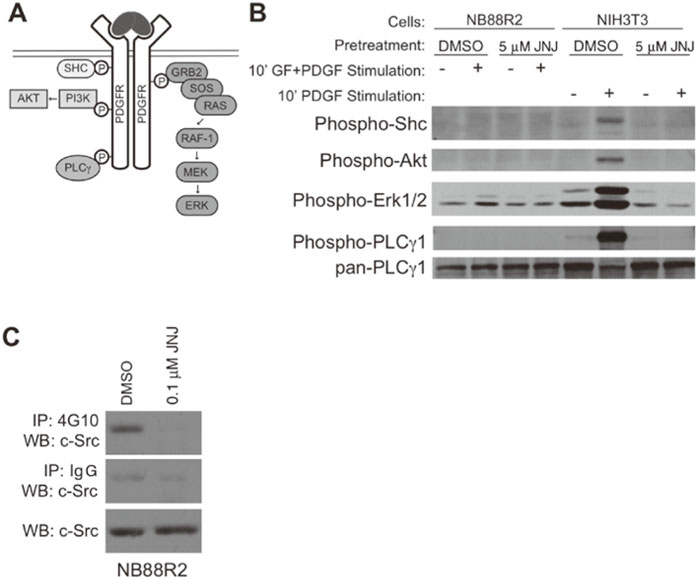
Fig. 3
JNJ-10198409 inhibits Src phosphorylation but does not alter PDGFR-dependent signal transduction in cells from NB patients.
A, Diagram of PDGF-dependent signal transduction pathways with selected PDGFR targets and interaction partners indicated.
B, NB88R2 or NIH-3T3 cells were pretreated with JNJ-10198409 and transient stimulation with PDGF as described in Methods. Immunoblots of NB88R2 cell lysates show no activation of PDGF-dependent signaling pathways while those for NIH-3T3 demonstrate PDGF-dependent stimulation of Shc, Akt, PLCγ and ERK1/2 phosphorylation that is blocked by JNJ-10198409 pretreatment of cells.
C, JNJ-10198409 treatment decreased c-Src phosphorylation in NB88R2 cells. Cells were treated with 0.1 μM JNJ-10198409 or DMSO for 48 hours, lysed, immunoprecipitated with anti-phosphotyrosine antibodies(4G10, top panel) or control IgG(middle panel) and immunoblotted for c-Src. Equal volume of input lysate was immunoblotted for c-Src(lower panel)
In this study, we characterized the effect of JNJ-10198409 on NB cell lines and primary cells from NB patients in vitro. JNJ-10198409, developed as a selective PDGFRβ kinase inhibitor, was identified in a cell-based screen for NB cell cytotoxic agents from a collection of 143 kinase inhibitors[2]. We demonstrated that treatment with nanomolar doses of JNJ-10198409 reduced the survival of NB cells from multiple patients and induced apoptotic cell death.
JNJ-10198409 was developed as an anti-angiogenic compound that also possessed modest anti-proliferative activity against a number of human cancer cell lines in vitro and in vivo[3,4]. JNJ-10198409 is an ATPcompetitive inhibitor of PDGFRβ with an IC50 of 4.2 nM in in vitro kinase assays[3]. To investigate the mechanism of JNJ-10198409 action in cells from NB patients, we first examined PDGF-dependent signaling. However, we did not observe activation of PDGFR downstream targets following transient PDGF stimulation. Additional kinase inhibitors present in the kinase-targeted chemical library with activity against PDGFR(AG 1296, mastinib/AB1010 and SU6668)were not identified as cytotoxic agents for cells from NB patients. Taken together, this data suggests that PDGFR is not the relevant target of JNJ-10198409 in those cells.
JNJ-10198409 is known to suppress the activities of c-Abl and the Src family kinases c-Src, Lck and Fyn in vitro[3], suggesting that these kinases may be the relevant targets for this compound. Previous studies have demonstrated that imatinib, which inhibits c-Abl, c-Kit and PDGFR kinase activities, decreased the proliferation of established NB cell lines in vitro[2,9-12]. However, the imatinib concentration required to inhibit NB cell line proliferation was significantly greater than that required to inhibit the growth of chronic myeloid leukemia cell lines which express an activated Abl oncogene[13]. In addition, various results were observed among NB xenograft tumor models[2,9,11,14]and no response was seen in a Phase II trial of refractory or recurrent NB patients[15]. In our laboratory, additional c-Abl-selective inhibitors present in the kinase-targeted chemical library were not identified as NB cell line hits. These observations suggest that c-Abl is also not the target of JNJ-10198409 in NB cells.
Of the known targets of JNJ-10198409, Src family kinases are the most likely candidates. A dual Src/c-Abl inhibitor, dasatinib, was reported to inhibit the proliferation of NB cell lines[16,17]and reduced tumor volume in NB xenograft tumor models[17]. We show here that treatment with nanomolar concentrations of JNJ-10198409 substantially decreased c-Src tyrosine phosphorylation in cells from NB patients, suggesting that Src family kinases are the therapeutically relevant target of JNJ-10198409.
Src family kinases are rarely mutated in human cancers but their expression or activity is frequently increased(reviewed in[18]). Despite the prevalence of Src activation in human cancers, targeted therapy against Src family kinases has only recently entered the clinic[19]. It is possible that JNJ-10198409 can be combined with other NB therapeutic agents and future studies will explore these combination therapies.
We thank L. Podmore and S. Mouaaz for assistance with sphere-formation experiments and L. Hansford and M. Fujitani for helpful discussions. This work was supported by the National Cancer Institute of Canada, The James Birrell Fund for Neuroblastoma Research, Lilah’s Fund, McLaughlin Centre for Molecular Medicine, and the Stem Cell Network. DRK holds a Canada Research Chair.
MF, NG, KMS and DRK conceived and designed the experiments. MF and NG performed the experiments. MF and KMS analyzed the data. MF and KMS co-wrote the paper, and DRK contributed revisions.
Abbreviations:NB; neuroblastoma, SKPs; skin-derived precursor cells
Address correspondence to Dr. Mayumi Fujitani.
Cell Biology Program and James Birrell Laboratories, The Hospital for Sick Children MaRS Centre, Toronto Medical Discovery Tower, 101 College Street, Room 12-314 M5G 1L7.
Phone: +1-416-813-7654(x 1432). Fax: +1-416-813-2212.
E-mail: fujimayu@msn.com
