




Chiba Medical J. 88E:35~39,2012
doi:10.20776/S03035476-88E-3-P35
[Original Paper]
Tadashi Hoshino 1,2), Naruhiko Ishiwada 1,3) and Yoichi Kohno 1)
1) Department of Pediatrics, Graduate School of Medicine, Chiba University, Chiba 260-8670.
2) Division of Infectious Diseases, Chiba Children’s Hospital, Chiba 266-0007.
3) Division of Control and Treatment of Infectious Diseases, Chiba University Hospital, Chiba 260-8677.
(Received April 5, 2012, Accepted April 18, 2012)
During the 10 years preceding the introduction of the Haemophilus influenzae type b(Hib) vaccine, restriction fragment length polymorphism(RFLP) analysis using pulsed-field gel electrophoresis was performed on 66 strains(22 sets) of Hib isolated simultaneously from cerebrospinal fluid, blood, and nasopharyngeal samples of 22 patients with meningitis. Strains from the 3 types of samples showed identical RFLP patterns in 18 of the 22 patients, confirming the pathway of Hib infection starting from the nasopharynx and leading to meningitis through bacteremia. In the remaining 4 patients, the RFLP patterns from the 3 sample types were also nearly identical. However, a single band in the RFLP patterns of strains from 1 of the 3 sample types showed a shift of about 20 kb. One copy of the cap-b gene had a molecular weight of 18 kb, and the discrepancy in the band’s molecular weight indicated an amplification or loss of a single copy of the cap. These findings suggest that Hib caused meningitis by changing the number of copies of the cap to evade the host’s immunological response.
Haemophilus influenzae type b, PFGE, RFLP, cap
In 2008, a Haemophilus influenzae type b(Hib) conjugate vaccine was introduced in Japan. However, even in 2011, Hib remains the major causative organism of bacterial meningitis in Japanese children because it is vaccinated voluntarily. Bacterial meningitis develops when bacteria carried in the nasopharynx spread, causing bacteremia, and then cross the blood-brain barrie[1]. Therefore, bacterial strains isolated from the nasopharynx, blood, and cerebrospinal fluid samples from patients with meningitis should be identical; however, few studies have reported the genetic homology between them[2]. In this study, we confirmed the pathway of development of bacterial meningitis in children by performing a restriction fragment length polymorphism(RFLP) analysis using pulsed-field gel electrophoresis(PFGE) of Hib isolates from the nasopharynx, blood, and cerebrospinal fluid ofpatients with meningitis.
This study was conducted on 38 patients with Hib meningitis who were admitted to Chiba Children’s Hospital and Chiba University Hospital during the 10-year period before the introduction of the Hib vaccine from 1998 to 2007. Before initiation of antimicrobial therapy, cerebrospinal fluid, blood, and/or nasopharyngeal samples were collected from patients in order to perform bacterial culture. The 22 patients from whom Hib was isolated in all 3 sample types were selected as study participants(Table 1). Other 16 patients were excluded because only 2 samples, CSF and blood, were collected. A slide agglutination test was performed at the time of isolation, and identification using an antiserum(DENKA SEIKEN Co., Ltd.)confirmed that the 66 bacterial strains from the 22 sets of samples were serotyped as b. The bacterial strains were stored at -80°C, and subcultures of the strain were kept to a minimum in order to prevent capsular loss.
Table 1
Patient number, year, age, sex, and criteria for RFLP patterns of 22 pediatric patients with purulent meningitis caused by H. influenzae type b.
RFLP analysis was performed using the PFGE method[3]. Bacterial genomic DNA was digested using the restriction enzyme SmaI(or ApaI for some samples)and was embedded in 1% agarose gels. Electrophoresis was then performed in 0.5×TBE buffer with run conditions of 170 V for 22 h at 14°C and pulse times of 5-50 s(for SmaI) or 2-25 s(for ApaI), ramped linearly using the CHEF-DR II system(Bio-Rad Laboratories, Hercules, CA, USA)
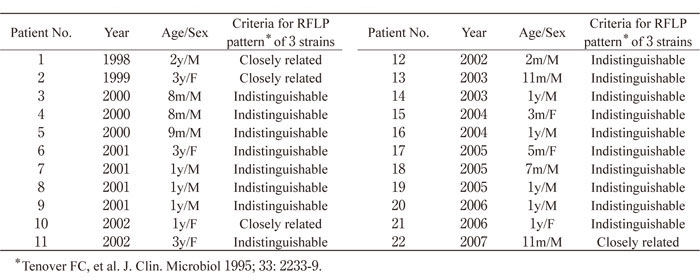
Figure 1 shows the RFLP patterns of the bacterial strains from the cerebrospinal fluid, blood, and nasopharynx by SmaI digested PFGE. In 18 of the 22 patients(patients 3-9 and 11-21), the RFLP patterns from the 3 types of samples were identical(Fig.1-a). For the remaining 4 patients(patients 1, 2, 10, and 22), bacterial strains from one of the samples showed a shift in molecular weight of about 20 kb for 1 band in the electrophoresis pattern(Fig.1-b). Figure 2 shows the electrophoresis results of bacterial strains from the 4 patients whose RFLP patterns after SmaI digested PFGE showed differences. In all 4 patients, the band with the different electrophoretic mobility had a molecular weight of about 485 kb. For patient 1, this band showed a shift of +20 kb in the strain isolated from the blood sample compared to those from the 2 other specimens. For patient 2, the band showed a shift of -20 kb in the cerebrospinal fluid isolates, and for patients 10 and 22, a shift of -20 kb was found in the nasopharyngeal isolates. The electrophoretic patterns of the strains isolated from those 4 patients were also compared after ApaI digestion(Fig.3). Differences were found in a band of about 120 kb. As with SmaI digestion, the band showed a shift in molecular weight of +20 kb in blood isolates from patient 1, a shift of -20 kb in the cerebrospinal fluid isolate from patient 2, and a shift of -20 kb in the nasopharyngeal isolate from patients 10 and 22.
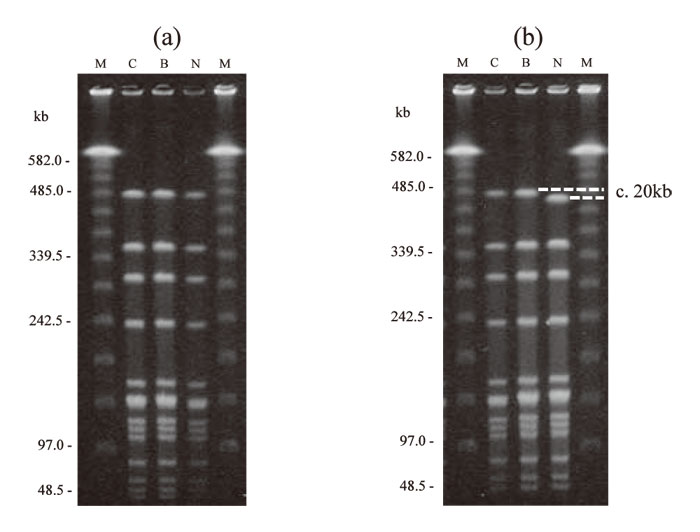
Fig.1
Examples of RFLP patterns of SmaI digested DNA of H. influenzae type b strains from patients with purulent meningitis. In 18 patients(patients 3-9 and 11-21), RFLP patterns of the 3 isolates were identical(Fig. 1-a). In 4 patients(patients 1, 2, 10, and 22), the patterns were identical except 1 strain in which a single band shift of approximately 20 kb was identified(Fig. 1-b).
Lanes:
C, isolate from cerebrospinal fluid;
B, from blood;
N, from nasopharynx;
M, lambda ladder molecular size marker.
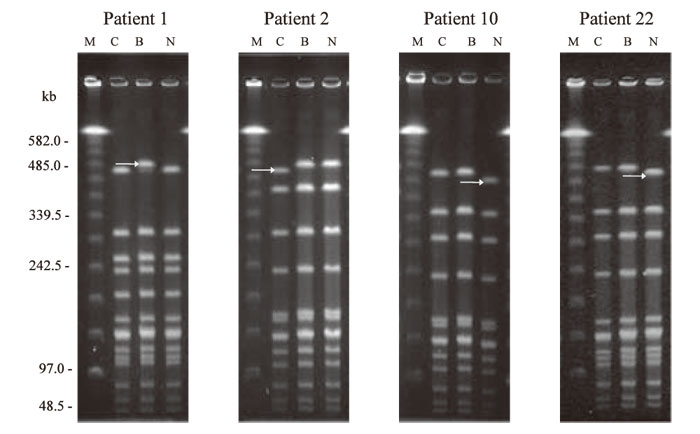
Fig.2
Discrepancy of RFLP patterns of SmaI digested DNA between isolates from cerebrospinal fluid, blood, and nasopharynx of 4 patients. In all patients, a difference in the ~ 485kb fragment was observed in 1 of 3 strains(arrow).
Patient 1, the isolate from the blood showed a singleband shift of approximately +20kb;
patient 2, the strain from the cerebrospinal fluid showed a shift of approximately -20kb;
patients 10 and 22, strains from the nasopharynx showed a shift of approximately -20kb shift.
Lanes:
C, isolate from cerebrospinal fluid;
B, from blood;
N, from nasopharynx;
M, lambda ladder molecular size marker.
Fig.3
Discrepancy of RFLP patterns of ApaI digested DNA between isolates from cerebrospinal fluid, blood, and nasopharynx of 4 patients. In all patients, a difference in the ~ 120kb fragment was observed in 1 of 3 strains(arrow).
Patient 1, the isolate from the blood showed a singlebandshift of approximately +20kb;
patient 2, the strain from the cerebrospinal fluid showed a shift of approximately -20kb;
patients 10 and 22, strains from the nasopharynx showed a shift of approximately -20kb shift.
Lanes:
C, isolate from cerebrospinal fluid;
B, from blood;
N, from nasopharynx;
M, lambda ladder molecular size marker.
In this study, isolates from cerebrospinal fluid, blood, and nasopharynx showed identical RFLP patterns in 18 of the 22 patients with Hib meningitis. For the remaining 4 patients, one of the bands had a slight difference in only 1 of the 3 sample types. The strains isolated from the 3 types of samples collected from the 18 patients that showed identical RFLP patterns were classified as“ indistinguishable,” and those from the 4 patients that showed a single band shift in the RFLP patterns were classified as“ closely related” interpreted using the criteria proposed by Tenover et al[4] (Table 1). As such, in patients whose RFLP patterns showed differences, the strains from the 3 different specimen types were also considered identical in nature. These findings demonstrate a pathway whereby bacterial strains carried in the nasopharynx led to the development of meningitis through bacteremia.
In all 4 patients, the size difference between the bands was about 20 kb. The expression level of the capsule(cap), which is the main virulence factor in capsulated H. influenzae strains, is regulated by the cap gene, which has a molecular weight of about 18 kb. Although Hib normally has 2 copies of the cap, this number of copies can change easily[5]. The difference in molecular weight was equivalent to the molecular weight of a single copy of the cap, suggesting that an amplification or deletion of the cap gene might have occurred between the isolates from the 3 types of samples. Similar findings have previously been reported about H. influenzae type a meningitis[6]. In that report, the RFLP pattern by PFGE revealed a band with a shift of 35 kb, that corresponded 2 copies of caps, between isolates from cerebrospinal fluid and blood. The quantitative real-time PCR analysis revealed that isolate from blood lost 2 copies of caps[6].
In 3 of the 4 patients(patients 1, 10, and 22), a band of approximately 485 kb in SmaI digested RFLP pattern of blood isolates showed a shift of about +20 kb compared to that in nasopharyngeal isolates. A similar shift was also found in a band of approximately 120 kb in ApaI digested RFLP pattern. Thus, it is likely that a single copy of the cap had been added. Hib has been reported to increase its resistance to complementmediated opsonization by increasing the number of copies of the cap[7]; likewise, our findings suggest that the number of copies of the cap in Hib might have increased upon invading the bloodstream in 3 patients. In addition, for 2 patients(patients 1 and 2), a band of ~485 kb in SmaI digested RFLP pattern of the cerebrospinal fluid isolates and a band of ~120 kb in ApaI digested RFLP pattern showed a shift of about -20 kb compared to that of blood isolates, suggesting a loss of one copy of the cap. In a previous study conducted on patients with vaccine failure, the number of copies of the cap was lower in isolates from patients with meningitis than in those with other systemic infections[8]. This may be because the thickness of the capsule impairs the expression of surface factors binding to cerebrovascular endothelial cells[8]. Accordingly, in the case of these 2 patients, the loss of the cap occurred when the Hib reached the subarachnoid space after having passed from the blood into the brain.
In this study, the number of copies was not confirmed by Southern blot[5]or quantitative real-time PCR[6]. However, the differences in RFLP patterns suggested that during the development of meningitis, Hib changed the number of copies of the cap to evade the host’s immunological response. There was no difference of clinical features between the patients in whom the RFLP patterns from 3 types of samples showed identical and those in whom the RFLP patterns were not same. However, changes in the number of copies of the cap have suggested to be associated with vaccine failure. According to previous reports, the incidence of strains with 3 or more copies of the cap is significantly higher in patients with vaccine failure, and the odds ratio of invasive Hib disease increases with the number of copies[8]. Although it has only been a short time since the Hib vaccine was introduced in Japan, Hib strains with more than 3 copies of the cap had been found in 16.7% of isolates from patients with meningitis even before the introduction of the vaccine[9]. The occurrence of vaccine failure will require attention in the future.
This manuscript has not been submitted or accepted for publication elsewhere.
Address correspondence to Dr. Tadashi Hoshino.
Division of Infectious diseases, Chiba Children’s Hospital, 579-1
Heta cho, Midori-ku, Chiba-City 266-0007, Japan.
Phone: +81-43-292-2111. Fax: +81-43-292-3815.
E-mail: t.hshn12@pref.chiba.lg.jp
