




Chiba Medical J. 88E:47~53,2012
doi:10.20776/S03035476-88E-4-P47
[Original Paper]
Harik Firman Thahadian 1) and Masahiko Hatano 1,2)
1) Department of Biomedical Science( M14), Graduate School of Medicine, Chiba University, Chiba 260-8670.
2) Department of Biomedical Science( M14), Graduate School of Medicine, Chiba University, Chiba 260-8670.
(Received December 15, 2011, Accepted April 6, 2012)
The Nczf gene identified as a target gene of Ncx encodes a novel Krab zinc finger protein which functions as a sequence specific transcriptional repressor. We investigated a role of Nczf in regulation of apoptosis. We stimulated NIH3T3 cells or murine thymocytes with various agents to induce apoptosis and examined the expression of Nczf mRNA. In NIH3T3 cells, Nczf mRNA expression was induced after X-ray irradiation or stimulation with adriamycin but not after UV irradiation or H2O2 stimulation. In thymocytes, the mRNA was induced after X-ray irradiation or dexamethasone stimulation. Furthermore, knockdown of Nczf in NIH3T3 cells resulted in increased apoptosis in culture. These results suggest that Nczf plays a protective role in regulation of apoptosis.
Nczf, Krab-zinc finger, Ncx, apoptosis, knock down
Nczf(Ncx regulated zinc finger) was identified as one of the novel target genes for Ncx. Nczf contains an N-terminal KRAB box domain and 11 Krüppel C2H2 type zinc finger motifs at C terminus[1]. KRAB box domain is responsible for protein-protein interaction and reported to recruit histone deacetylase complex[2]. Zinc finger domain is responsible for sequence specific DNA binding[3]. Nczf localized in the nucleus and functioned as a transcriptional repressor. Promoter region of the Nczf gene contains 4 consensus sequences for Ncx-binding motifs. Transient transfection assays of the 5’-flanking region fused to the luciferase reporter gene with various amounts of the Ncx expression vector showed a dose dependent increase of the Nczf promoter activity.
The murine Ncx(Tlx2 , Enx, Hox11 L1) gene is specifically expressed in neural crest derived tissues and regulates neuronal cell death in enteric neurons[4]. Mice deficient for Ncx developed megacolon with the increased number of enteric neurons[5,6]. The pathology of Ncx-deficient mice resembles intestinal neuronal dysplasia in humans[5-8]. The expression of Nczf and Ncx mRNA was detected in enteric neurons. The amount of Nczf mRNA in enteric ganglia from Ncx deficient mice decreased to one third of wild type ones.
Although the expression of Ncx is restricted to neural crest derived tissues, Nczf mRNA is ubiquitously expressed in various adult mice tissues. This fact suggests that Nczf expression is regulated not only by Ncx in neural crest derived cells but also by other factors in various situations. Since Ncx regulates cell death of enteric neurons[9], it is possible that its target genes also regulate cell death in various tissues and in various situations.
Apoptosis is the most studied form of programmed cell death. Apoptosis plays a fundamental role in normal development and tissue homeostasis. Abnormal regulation of apoptosis is associated with variety of diseases such as immunological and developmental abnormalities, neurodegeneration, and cancer[10]. Genetic studies of C. elegans led to the identification of important molecules for apoptosis regulation, which were conserved evolutionally. The subsequent characterization of the corresponding genes elucidated the basic molecular machinery of apoptosis[11,12]. However, involvement of Nczf in apoptosis signaling pathway is not studied.
In this report, we examined the expression of Nczf mRNA in apoptosis using NIH3T3 cells and murine thymocytes. Nczf mRNA was induced by various stimuli of apoptosis including X-ray irradiation, adriamycin and dexamethasone. Furthermore, knock down of Nczf in NIH3T3 cells resulted in increased apoptosis in culture. We will discuss the possible role of Nczf in apoptosis.
Cell culture and stimulation NIH3T3 cells were maintained in Dulbecco’s modified Eagle’s medium(DMEM)(Sigma) supplemented with 10% fetal calf serum(FCS)(Sigma) at 37℃ . Thymocytes were prepared from C57BL/6 mice(Japan Clea Co. LTD) and cultured in RPMI1640 medium supplemented with 10% FCS. Those cells were plated on a 60mm dish at 1.5 x 105 cells per each dish 1 day before stimulation. For X-ray irradiation, cells were collected in 14ml plastic tube, suspended in PBS and exposed to X-ray using HITEX cabinet X-ray system® (150kV, 10mA with 1.0mm Al and 0.1mm Cu filters) (10Gy for NIH3T3 cells and 2Gy for thymocytes). For ultraviolet exposure, attached cells were washed 3 times with PBS on a 60mm tissue culture dish, covered with 1ml of PBS and exposed to ultraviolet(100 J/M2 in FUNA-UV-LINKER FS-800® , 254nm). In some experiments, Adriamycin(200ng/ml), H2O2(200μM) or dexamethasone(100nM) were added to the culture.
For knockdown experiments, shRNA for Nczf (5’-GTA TTG TAG AGC GAT ATAA-3’) and scrambled control RNA(5’-TAG AGT GTT GAT CAG AATA- 3’) were designed by TAKARA and inserted into RNAi- Ready pSIREN-RetroQ ZsGreen siRNA expression vector(TAKARA). Lipofection was performed using FuGENE 6 transfection kit(Roche Applied Science) according to the manufacturer’s recommendation.
All animal experimental procedures were approved by the Institutional Animal Care and Use Committee at Chiba University.
Quantitative real-time PCR Total RNA was prepared by using TRIzol(Invitrogen) according to the manufacturer’s protocol. Each total RNA sample was then reverse-transcribed using a SuperScript first-strand synthesis system for RT-PCR(Invitrogen) according to the manufacturer's instructions. Quantitative Real-time PCR with the cDNA was conducted using a SYBR Green PCR Master Mix(PE Applied Biosystems) and run on an ABI Prism 7000 Sequence Detection System(PE Applied Biosystems). Primers were as follows:
Nczf F: 5’-CAG AAT CTC AAC GAC GCT CAG A-3’
Nczf R: 5’-AAC TCA GGT TTA GGG ATG CAT TG-3’
β-actin F: 5’-CCA GCC TTC CTT CTT GGG TAT-3’
β-actin R: 5’-TGG CAT AGA GGT CTT TAC GGA TGT-3’.
Data were analyzed using Sequence Detector software (PE Applied Biosystems). In addition to duplicate reactions for gene expression in each sample, each real-time PCR plate contained reactions for generating standard curves using serial dilutions of a known quantity of cDNA. The data were reported as the ratio of the calculated amount of candidate RNA in a given sample by the calculated amount of the housekeeping control(β-actin) gene in the same sample.
Apoptotic analysis Apoptotic cells were detected using an Annexin V Apoptosis detection kit 1 following the manufacturer’s instructions(BD Pharmingen). Annexin V positive apoptotic cells were analyzed on FACS Calibur (BD Biosciences) using Cell Quest software(BD Biosciences).
Statistical analysis Data were analyzed using a single-tailed Student’s t test. Data are given as mean ± S.D.
Nczf expression is induced by X-ray irradiation In order to know the possible participation of Nczf in cell death, we examined the expression of Nczf in cultured and primary cells after various stimuli. NIH3T3 cells were exposed to X-ray(10Gy) and cultured up to 72 hours. The cells were harvested every 24 hours and apoptotic cells were examined by flow cytometer. After 72 hours, percentage of apoptotic cells increased up to 20%. The expression of Nczf mRNA increased more than 2 folds 72 hours after X-ray irradiation(Fig.1A). Thymocytes are sensitive to X-ray irradiation and undergo apoptotic cell death. We exposed murine thymocytes to 2 Gy of X-ray and examined the apoptosis and the Nczf expression. The percentage of apoptotic cells increased after irradiation. The expression of Nczf mRNA also increased up to ten folds 10 hours after irradiation(Fig.1B). These data suggest that Nczf participates in cellular process of apoptosis induced by X-ray irradiation
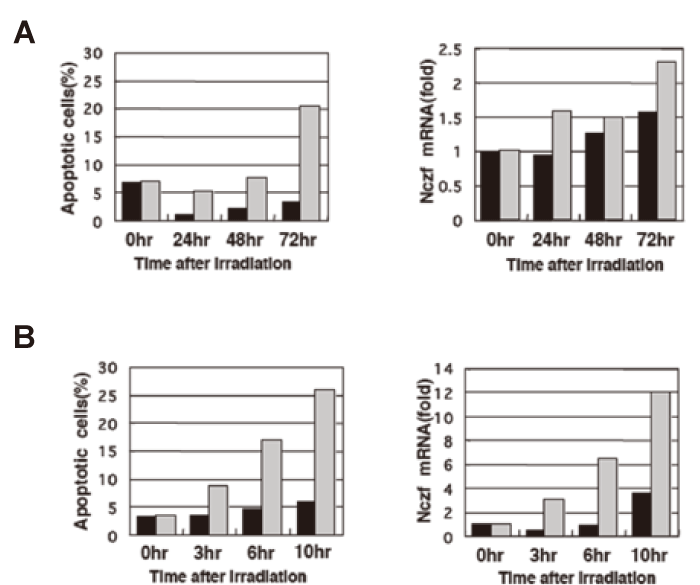
Fig.1
Expression of Nczf after induction of apoptosis by X-ray irradiation:
(A) NIH3T3 cells were exposed to 10Gy of X-ray irradiation and cultured in vitro. Cells were harvested at the indicated time and percentage of apoptotic cells were examined by FACS(left panel). RNA was isolated at each time point and Nczf mRNA expression was examined by real time PCR (right panel). The data are expressed as relative to the level of control(0hr without irradiation). Closed bar: without X-ray irradiation, shaded bar: with X-ray irradiation.
(B) Murine thymocytes were exposed to 2Gy of X-ray irradiation and cultured in vitro. Cells were harvested at the indicated time and percentage of apoptotic cells was examined by FACS(left panel). RNA was isolated at each time point and Nczf mRNA expression was examined by real time PCR(right panel). The data are expressed as relative to the level of control(0hr without irradiation). Closed bar: without X-ray irradiation, shaded bar: with X-ray irradiation.
Nczf expression is induced by adriamycin or dexamethasone induced cell death To further examine the relationship between Nczf and cell death, adriamycin or dezamethasone was used to induce apoptosis on NIH3T3 cells or murine thymocytes, respectively. When NIH3T3 cells were cultured in the presence of 200ng/ml of adriamycin, the percentage of Annexin V positive apoptotic cells increased(Fig.2A). Expression of Nczf mRNA increased more than 2-fold and the expression level correlated to the percentage of apoptotic cells(Fig.2B).
Dexamethasone induces apoptosis in murine thymocytes. We cultured thymocytes in the presence of 100nM dezamethasone for 5 hours and examined the percentage of apoptotic cells by flow cytometer. As shown in Fig. 3A, more than 25% of the thymocytes cultured in the presence of dexamethasone underwent apoptosis. The expression of Nczf mRNA from the thymocytes exposed to dexamethasone was 9-fold higher than that from freshly isolated thymocytes(Fig.3B). Nczf mRNA expression was also induced 5-fold in thymocytes cultured without dexamethasone for 5 hours, probably due to spontaneous apoptosis of thymocytes.
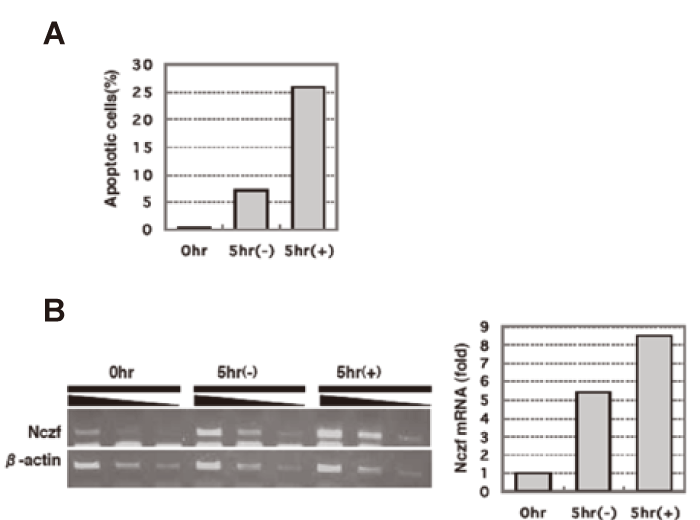
Fig.2
Expression of Nczf afterinduction of apoptosis by adriamycin:
(A) NIH3T3 cells were cultured in the presence of adriamycin(200ng/ml). Cells were harvested at the indicated time and percentage of apoptotic cells were examined by FACS(left panel). Percentage of apoptotic cells (AnnexinV+PI-cells) was plotted(right panel). Closed bar: without adriamycin, shaded bar: with adriamycin.
(B) RNA was isolated at each time point and Nczf mRNA expression was examined by reverse transcribed PCR(left panel) or qPCR (right panel). The data are expressed as relative to the level of control(0hr). Closed bar: without adriamycin, shaded bar: with adriamycin.
Fig.3
Expression of Nczf after induction of apoptosis by dexamethasone:
(A) Murine thymocytes were cultured in the presence or absence of dexamethasone(100nM) for 5 hours and cells were collected. Percentage of apoptotic cells was examined by FACS.
(B) RNA was isolated at each time point and Nczf mRNA expression was examined by reverse transcribed PCR(left panel) or qPCR(right panel). The data are expressed as relative to the level of control(0hr). In(A) and (B),(-) indicates without dexamethasone and (+) indicates with dexamethasone.
Nczf expression is not induced by H2O2 or ultraviolet induced cell death Hydrogen peroxide(H2O2) induces apoptosis by oxdative stress. When NIH3T3 cells were cultured in the presence of 200μM of H2O2, more than 10% of the cells underwent apoptosis(Fig.4A). The expression of Nczf mRNA did not increase in H2O2 induced apoptosis in NIH3T3 cells. Exposure of cells to ultraviolet(UV) induces DNA damage and causes apoptosis. We exposed NIH3T3 cells to UV(100J/m2) and induced apoptosis(Fig.4B). Again, Nczf mRNA expression did not increase in UV induced apoptosis(Fig.4B). These data suggest that induction of Nczf mRNA is not a general event associated with apoptosis.
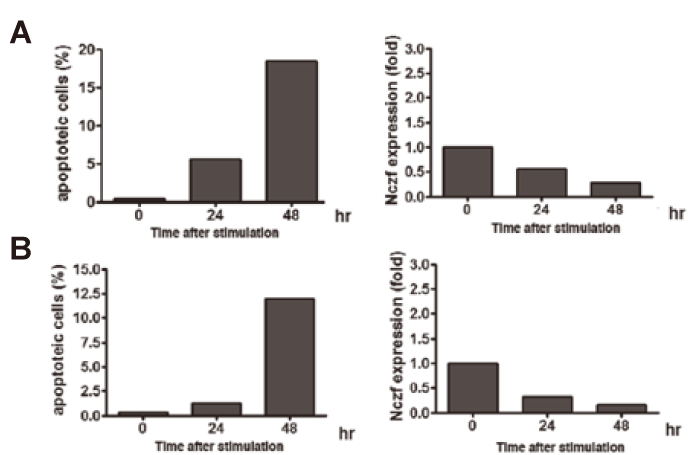
Fig.4
Expression of Nczf after induction of apoptosis by H2O2 or ultraviolet irradiation:
(A) NIH3T3 cells were cultured in the presence of H2O2. Cells were harvested at the indicated time and percentage of apoptotic cells were examined by FACS(left panel). RNA was isolated at each time point and Nczf mRNA expression was examined by real time PCR(right panel). The data are expressed as relative to the level of control(0hr).
(B) NIH3T3 cells were exposed to 100J/m2 of UV and cultured in vitro. Cells were harvested at the indicated time and percentage of apoptotic cells were examined by FACS(left panel). RNA was isolated at each time point and Nczf mRNA expression was examined by real time quantitative PCR(right panel). The data are expressed as relative to the level of control(0hr)
Knock down of Nczf in NIH3T3 cells results in increased susceptibility to apoptosis In order to elucidate the role of Nczf in apoptosis, we knocked down the expression of Nczf using shRNA system in NIH3T3 cells. We constructed bicistronic vectors expressing both a target gene and green fluorescent protein(GFP) to create a system in which the effect of an siRNA sequence was reflected in the GFP expression level(Nczf shRNA-pSIREN ZsGreen and scrambled shRNA-pSIREN ZsGreen as a control). Transfection of Nczf shRNA expression vector reduced the expression of Nczf to 50%(Fig. 5A, GFP+cells). The cells were maintained in DMED supplemented with 10% FCS. Percentage of apoptotic cells was examined in each population every 24 hours. As shown in Fig. 5B, Nczf shRNA transfected knock down cells(GFP+ population) were more susceptible to apoptosis in normal culture condition. These data suggest that Nczf protects cells from apoptosis induced by some cellular stress during in vitro culture.
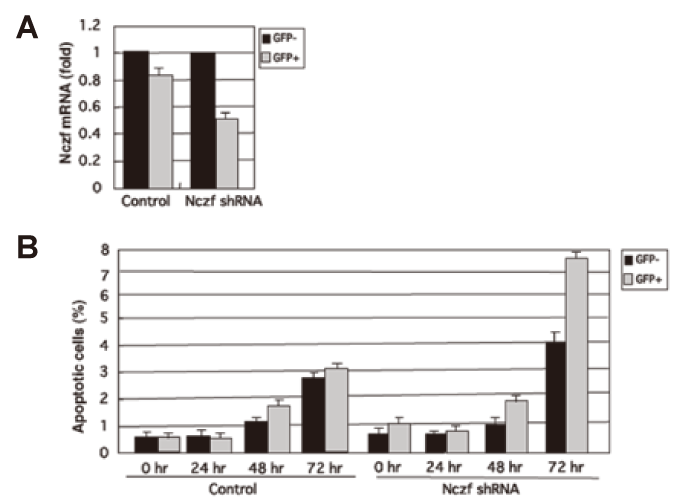
Fig.5
Knock down of Nczf in NIH3T3 cells:
(A) Expression of Nczf mRNA in NIH3T3 cells transfected with scrambled shRNA-pSIREN ZsGreen(control) or Nczf shRNA-pSIREN ZsGreen(Nczf shRNA) vectors. Cells were collected 24hour after transfection and GFP+ and GFP- cells were separated by FACS. RNA was isolated from each population and Nczf mRNA expression was examined by real time quantitative PCR. The data are expressed as relative to the level of control(GFPpopulation). Closed bar: GFP- population, shaded bar: GFP+ population.
(B) NIH3T3 cells transfected with scrambled shRNA-pSIREN ZsGreen(control) or Nczf shRNA-pSIREN ZsGreen(Nczf shRNA) vectors were maintained in DMEM with 10% FCS. Cells were collected at indicated time points and percentage of apoptotic cells in GFP- and GFP+ populations were examined by FACS. Closed bar: percentage of apoptotic cells in GFP- population, shaded bar: percentage of apoptotic cells in GFP+ population.
Nczf encodes a Krab-zinc finger protein and is expressed ubiquitously in adult mouse tissues. We have demonstrated that Nczf expression increased in certain apoptotic stimuli. X-ray irradiation causes double strand DNA breaks and induces apoptosis. In response to DNA double strand breaks, expression of p53 is induced after X-ray irradiation. Then, G1 arrest of cell cycle takes place and DNA repair machinery is activated [13]. If the damage of genome is too large to repair, the cell undergoes apoptosis. Nczf expression is increased 2-fold in NIH3T3 cells and more than 10-fold in thymocytes after X-ray irradiation. Adriamycin, an anticancer agent, also induces DNA double strand breaks and p53 expression, and causes apoptosis[14]. Nczf expression is also induced after adriamycin treatment in NIH3T3 cells. Induction of p53 protein reaches maximum within 3 hours after DNA damage(data not shown). The timing of Nczf induction is later than that of p53 induction. It is possible that Nczf functions in downstream of the p53 pathway. Alternatively, Nczf may participate in regulation of cell cycle or DNA repair machinery.
Dexamethasone is the most effective antiinflammatory drugs and directly induces T cell apoptosis. But the precise mechanism is not fully understood. Dexamethasone binds to the glucocorticoid receptor and thymocytes respond to dexamethasone by Bax translocation to mitochondria, opening the mitochondrial permeability transition pore, followed by a loss of mitochondrial transmembrane electric potential, and by the release of cytochrome c[15-17]. During the process of dexamethasone-induced apoptosis in thymocytes, expression of Nczf mRNA increased more than 8-fold. Nczf expression also increased 5-fold without dexamethasone after 5 hours of culture. Immature thymocytes spontaneously undergo apoptosis in in vitro culture. Probably, the induction of Nczf without dexamethasone is explained by spontaneous apoptosis in vitro.
Induction of the Nczf mRNA is not a general event accompanied with apoptosis. H2O2 and UV induce apoptosis in NIH3T3 cells but neither H2O2 nor UV stimulation induced Nczf mRNA expression. Reactive oxygen species(ROS) such as H2O2 induces mitochondrial dysfunction with activation of caspases and results in apoptosis. Cells exposed to UV produce ROS and cause mitochondrial dysfunction[18]. UV also causes DNA damage by forming thymine dimers and activates p53 pathways[19]. At this moment, it is not clear which specific pathway of apoptosis is responsible for Nczf induction. In addition, it is possible that the amount of Nczf is regulated at protein level. Further analysis is required to elucidate the function of Nczf in apoptosis signaling pathway.
Finally, we tried to knock down the Nczf expression by shRNA in NIH3T3 cells. When we maintain the knock down cells, percentages of the apoptotic cells increase by day 3. We could not get efficient knock down clones. The efficiency of the mRNA reduction was at most ~50%. It is possible that further reduction of Nczf expression is disadvantageous to the cells. These data suggest that Nczf plays a role to protect cells from apoptosis induced by stress during cell proliferation. We tried to examine whether over-expression of Nczf could protect cells from apoptosis. Bicistronic Nczf and GFP expression vector was transfected to NIH3T3 cells. However, we could not get GFP positive Nczf expressing transfectants. Probably the amount of Nczf is strictly regulated in a narrow physiological range in each cell and excess amount of Nczf may be harmful. Given the fact that Nczf functions as a transcriptional repressor, it is possible that Nczf negatively regulates the expression of pro-apoptotic genes. We then examined the expression of Bax, Bak, p53 and caspase3 in the knock down cells. Expression of these genes did not change in the knock down cells(data not shown). Identification of Nczf target genes will reveal the molecular mechanism of apoptosis protection by Nczf. In conclusion, Nczf mRNA is induced in some situation of apoptosis signaling and may play a protective role in regulation of apoptosis.
We thank M. Hanazono and A. Goda for skillful technical assistance, Drs. L. Fujimura, H. Takano, Y. Teratake, C. Ozeki, Y. Hasegawa and T. Tokuhisa for helpful discussion. This work was supported by a Grantin- Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Abbreviations:GFP; green fluorescent protein, UV; ultraviolet
Address correspondence to Dr. Harik Firman Thahadian.
Department of Biomedical Science(M14), Graduate School of Medicine, Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba 260-8670, Japan.
Phone: +81-43-226-2950. Fax: +1-81-43-226-2953.
E-mail: hatanom@faculty.chiba-u.jp
