




Chiba Medical J. 89E:41~47,2013
doi:10.20776/S03035476-89E-6-P41
[The Chiba Medical Society Award (2013)]
Kotaro Suzuki
Department of Allergy and Clinical Immunology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
(Accepted July 17, 2013)
Mast cells are known to play a pivotal role in allergic diseases such as allergic rhinitis, asthma, and atopic dermatitis by releasing granules containing histamine, LTC4, and other preformed chemical mediators. However, the precise regulatory mechanisms that lead to IgE-dependent exocytosis in mast cells have been unknown. Recently, I have shown that IKK2 (also called IKKβ), a catalytic subunit of the IKK complex and a central component of the intracellular signaling pathway mediating NF-κB activation, induces IgE-mediated degranulation in mast cells by phosphorylating SNAP-23, the target membrane soluble N-ethylmaleimide-sensitive fusion factor attachment protein receptor (SNARE).
This review highlights the molecular mechanisms of IKK2-mediated degranulation in mast cells and provides a novel potential therapeutic approach for allergic diseases.
Mast cells, degranulation, IKK2, NF-κB, anaphylaxis, SNARE
Mast cells are recognized as the major effector cells of the type I hypersensitivity reactions because of their high-affinity receptors for IgE (FcɛRI). They are known to play a pivotal role in allergic diseases, such as atopic rhinitis, asthma, and atopic dermatitis. Engagement of FcɛRI by IgE, followed by the aggregation of multiple IgE-bearing FcɛRI molecules by polyvalent antigen, leads to degranulation, and release of histamine, LTC4 and other preformed chemical mediators. Additionally multiple cytokine genes are transcribed, newly synthesized arachidonic acid metabolites are secreted, all of which trigger allergic inflammation. Though it is well established that mast cells mediate the early phase of type I hypersensitivity reactions by releasing granule contents after FcɛRI crosslinking, the precise regulatory mechanisms that lead to IgE-dependent exocytosis in mast cells have been unknown.
SNARE (soluble N-ethylmaleimide-sensitive fusion factor attachment protein receptor) proteins play a central regulatory role in exocytosis by promoting vesicle fusion with the plasma membrane[1]. The association of v-SNAREs and t-SNAREs to form a ternary SNARE complex that catalyses membrane fusion is required for exocytotic processes in many cells (Fig. 1). The SNARE complex brings the two membranes in apposition, a necessary step in overcoming the energy barrier required for membrane fusion. In mast cells, these include syntaxin 4 and SNAP-23 as t-SNAREs, while VAMP-2 and VAMP-8 represent candidates for v-SNAREs. The syntaxin-binding protein Munc18b and Munc18c are also essential for exocytosis in mast cells. It has been shown that phosphorylation of SNAP- 23 modulates exocytosis[2]. Though protein kinase C can phosphorylate SNAP-23 in vitro, it has not been established as to which kinase phosphorylates SNAP-23 in vivo and regulates exocytosis in mast cells [2].
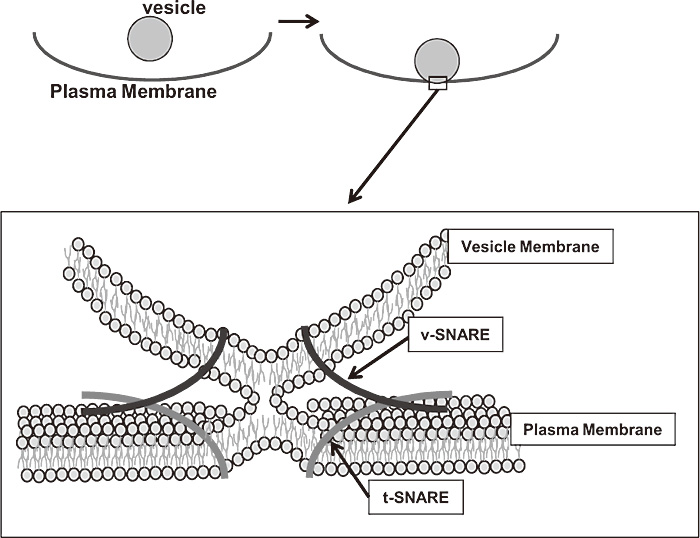
Fig. 1
SNARE pin formation is required for exocytotic process.
The association of v-SNAREs on the vesicle membrane and t-SNAREs on the plasma membrane to form a ternary SNARE complex that catalyses membrane fusion is required for exocytotic processes in many cells. In mast cells, the exocytotic processes is called“ degranulation”.
Transcription factors of nuclear factor-κB (NF-κB) is one of the major signaling pathways activated when cells are exposed to a variety of stimuli, including cytokines, such as TNF and IL-1, ultraviolet (UV) radiation, stress, and assaults. Upon activation, NF-κB regulate hundreds of genes in the context of multiple important biological processes, such as apoptosis, proliferation, innate and adaptive immune responses, and inflammation. The IKK-complex coordinates the response to most of the external signals leading to the induction of NF-κB activated gene transcription. The activity of the IKK-complex resides in two catalytic subunits, IKK1 (also called IKK α) and IKK2 (also called IKK β), and two regulatory subunits, NEMO and ELKs. Interestingly IKK2 is the major cytokineresponsive IκB kinase and thus is a central component of the intracellular signaling pathway mediating NF-κB activation. IKK2 shares sequence and structural similarity with IKK1 (50% sequence identity). The crystal structure analysis of IKK2 in the IKK complex reveals a trimodular architecture comprising the kinase domain, a ubiquitin-like domain, and an elongated, α-helical scaffold/dimerization domain. The ubiquitinlike domain and scaffold/dimerization domain mediate a critical interaction with IκBα that restricts substrate specificity. The ubiquitin-like domain is also required for catalytic activity of IKK2. The scaffold/dimerization domain mediates IKK2 dimerization that is required for IKK2 activation. IKK2 is responsible for the majority of IKK activity in most cell types. Consequently, IKK2- deficient cells or mice exhibit less phosphorylation of IκBα, resulting in defective NF-κB activation.
In addition to NF-κB-dependent functions of IKK2, accumulated evidences suggest that IKK2 regulates some biological phenomenon in a NF-κB-independent manner.
Roles of IKK2 and/or NF-kBs in mast cellmediated allergic reactions have been undetermined.
Recently, I have shown that IKK2 induces IgE-mediated degranulation in mast cells by phosphorylating SNAP- 23, the target membrane soluble N-ethylmaleimidesensitive fusion factor attachment protein receptor (SNARE), and that this novel IKK2-mediated function is completely NF-κB-independent [3].
To investigate the role of IKK2 on anaphylactic responses, which is induced by mast cells, I used mast cell knock-in mouse model [4]. Fetal liver-derived mast cells (FLMCs) were generated from days 11.5-12.5 embryonic liver of wild type (WT) or IKK2-deficient (IKK2-/-) mice because of the lethality in the IKK2-/- embryos at E14[5]. To generate mast cell knock-in mice, cultured FLMCs were transplanted intradermally (i.d.) or intravenously (i.v.) into mast cell-deficient WBB6F1-Kitw/Kitw-v (W/Wv) mice. Four weeks after transplantation, IgE-dependent anaphylactic reactions in mast cell-reconstituted mice were examined.
As shown in Figure 2(Fig. 2), antigen challenge of mice that had been sensitized previously with monoclonal DNPspecific IgE antibody demonstrated that IKK2-/- FLMCs reconstituted mice exhibited reduced cutaneous anaphylaxis than the WT FLMCs reconstituted mice, as assessed by change in ear. I next examined passive systemic anaphylaxis by measuring decrease of body temperature after antigen challenge. Figure 3(Fig. 3) shows a decrease in passive systemic anaphylaxis in the IKK2-/- FLMCs reconstituted mice. These data imply a physiological role of IKK2 in mast cells is to induce appropriate mast cell degranulation.
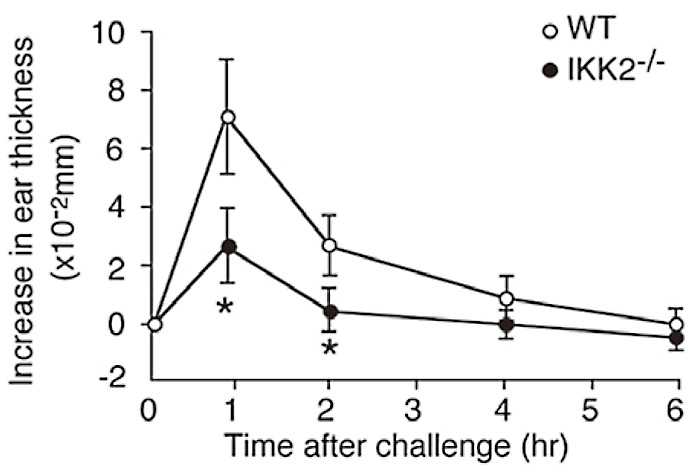
Fig. 2
Severe impairment of passive cutaneous anaphylaxis in the absence of IKK2.
For passive cutaneous anaphylaxis (PCA), WT or IKK2-/- mast cells were injected intradermally into the right ear of W/Wv mice. Four weeks later, mice were first primed with antigen-specific IgE by intradermal injection twenty-four hours later, the mice were injected with antigen. The degree of PCA was estimated by quantification of ear swelling. P<0.01.( ref.3)
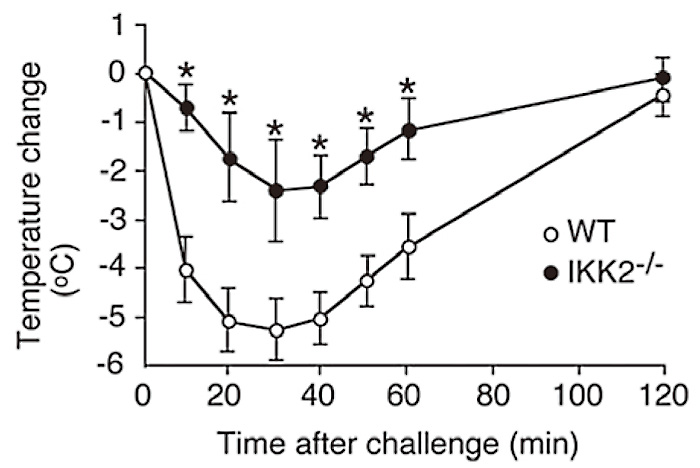
Fig. 3
Severe impairment of passive cutaneous anaphylaxis in the absence of IKK2.
For systemic anaphylaxis, we injected WT or IKK2-/-mast cells, into W/Wv mice to elicit systemic anaphylactic reactions. Four weeks later, mice were primed with antigen-specific IgE by i.v. injection. After another twenty-four hours, mice were injected with antigen and the systemic anaphylactic response was monitored by measuring rectal temperature. *P<0.01. (ref.3)
To investigate the cellular basis for the diminished in vivo anaphylactic reactions in IKK2-/- FLMCs reconstituted mice, I next compared degranulation (release of β-hexosaminidase and histamine which distribute in mast cell granule at steady state) between FLMCs derived from WT and IKK2-/- mice. Consistent with the defect in anaphylactic responses in vivo, IKK2-/- FLMCs showed clear defects in FcɛRIstimulated β-hexosaminidase release (Fig. 4A), and %histamine release (Fig. 4B). I also examined the effect of over-expression of IKK2 in the mast cells on FcɛRIstimulated β-hexosaminidase release. RBL-2H3, a rat basophilic cell line stably expressing GFP-IKK2 resulted in the increase of FcɛRI-stimulated β-hexosaminidase release (Fig. 5). To determine whether kinase activity of IKK2 is required for IKK2-mediated up-regulation of FcɛRI-stimulated β-hexosaminidase release, the effect of over-expression of IKK2KM, a kinase inactive mutant of IKK2, on FcɛRI-stimulated β-hexosaminidase release was examined. Over-expression of IKK2KM did not enhance FcɛRI-stimulated β-hexosaminidase release (Fig. 5). These results suggest that IKK2 modulates antigen-induced mast cell degranulation in a kinase activity-dependent manner.
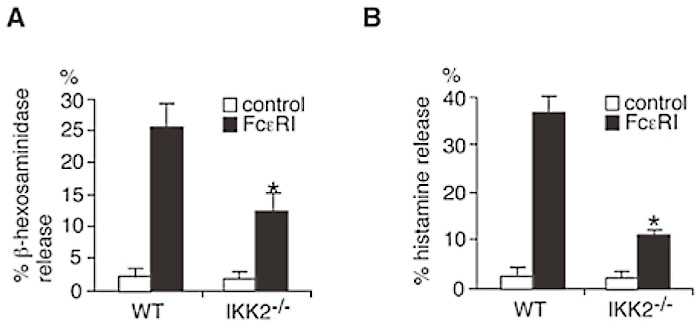
Fig. 4
IKK2 plays critical roles in mast cell degranulation.
(A) IgE-induced β-hexosaminidase release is impaired in IKK2-/- mast cells. WT or IKK2-/- mast cells were cross-linked upon FcɛRI. Thirty minutes later, β-hexosaminidase activity in the supernatants and lysate of the cell pelletes was quantified. *P<0.01.(B) IgEinduced histamine release is impaired in IKK2-/- mast cells. WT or IKK2-/- mast cells were cross-linked upon FcɛRI. Thirty minutes later, cell-free supernatants were recovered. Histamine was measured by EIA. *P<0.01. (ref.3)
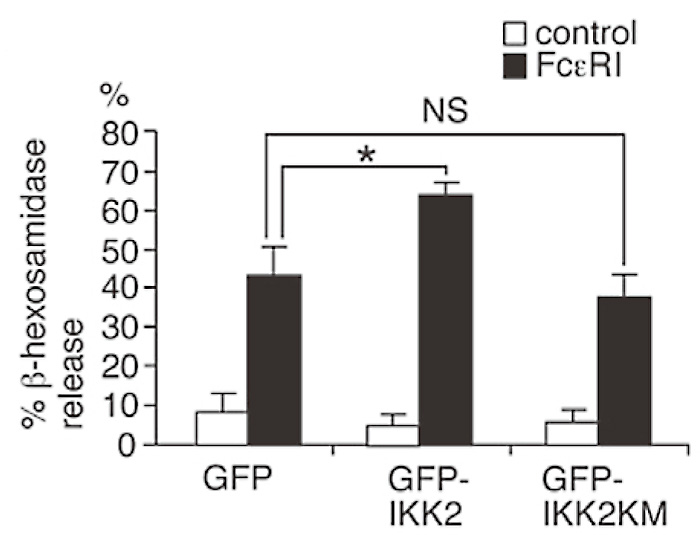
Fig. 5
IKK2 plays critical roles in mast cell degranulation.
RBL-2H3 cells (basophil line) were transfected with pcDNA3.1/Zeo GFP-IKK2, pcDNA3.1/Zeo GFPIKK2KM, and pcDNA3.1/Zeo GFP vector( as control).
Cells were cross-linked upon FcɛRI. Thirty minutes later, β-hexosaminidase activity in the supernatants and lysate of the cell pelletes was quantified. *P<0.01. NS, not significant. (ref.3)
The seminal event in NF-κB activation is the phosphorylation of IκBs, which is mediated by IKK complex. Therefore, the role of NF-κB activation in FcɛRI-induced degranulation was examined.
NF-κB activation and IgE-induced degranulation were examined in RBL-2H3 cells stably expressing IκBα superrepressor (IκBαM). As expected, expression of IκBαM in the RBL-2H3 cells resulted in the complete inhibition of NF-κB-dependent promoter activation in response to TNF alpha (Fig. 6A). In contrast, IκBαM-expressing RBL-2H3 cells showed similar levels of FcɛRI-stimulated β-hexosaminidase release to the control cells. (Fig. 6B). It thus appears that FcɛRI-induced degranulation is independent of NF-κB activation pathway. I thus conclude that IKK2 modulates IgE-induced degranulation by NF-κB-independent mechanism.
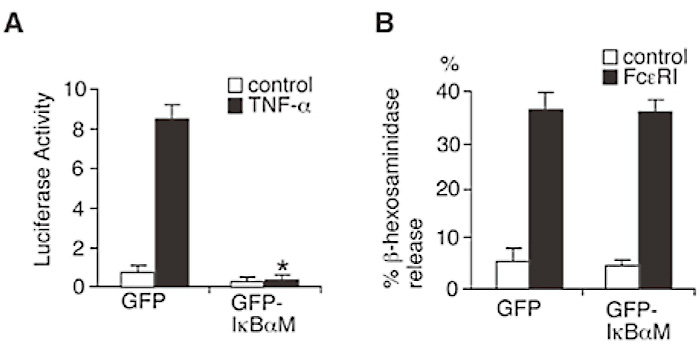
Fig. 6
NF-κB activation is not essential for mast cell degranulation.
(A) RBL-2H3 cells were transfected with pcDNA3.1/ Zeo IκBαM( dominant negative mutantof IkBα) and pcDNA3.1/Zeo mock vector( as control). Cells were transfected with HIV-3x-κB luciferase and β-galactosidase plasmid. 24 hours later, cells were stimulated with( black columns) or without( white columns) TNF-α. 6 hours later, luciferase activity was measured. *P<0.01.(B) Cells were cross-linked upon FceRI. Thirty minutes later, β-hexosaminidase activity in the supernatants and lysate of the cell pelletes was quantified. (ref.3)
Mast cells contain the molecular machinery that drives membrane fusion during granule exocytosis.
Essential to this process are SNARE [soluble NSF (N-ehyl-maleimide-sensitive factor) attachment protein receptor] proteins that lie on opposing cellular membranes to form a stable multimeric complex that catalyze fusion[1]. Mast cells express several plasma membrane-localized SNAREs (Q-SNAREs) that form part of the exocytotic machinery. Both SNAP-23 and syntaxin 4 have been implicated in IgE-dependent fusion and are partly localized in lipid rafts, domains of the plasma membrane enriched in sphingolipids, and cholesterol. Furthermore, it has been suggested that, in T cells, IKK1 and IKK2 are recruited to the lipid rafts at the immune synapse upon TCR activation[6], therefore I first examined the recruitment of IKK2 into the lipid rafts after FcɛRI-stimulation. SNAP-23 and syntaxin 4 were detected in the membrane raft fraction before and after FcɛRI stimulation (Fig. 7 (Fig. 7), co-sedimentation with CTx in fraction 5). Little or no IKK2 was detected in the lipid rafts. In contrast, substantially increased levels of IKK2 were detected in the membrane raft fraction upon FcɛRI stimulation (Fig. 7), suggesting that IKK2 co-localize with SNAP-23 and syntaxin 4 in the lipid rafts only after stimulation. Furthermore, I examined whether IKK2 associates with SNAP-23 in the lipid rafts before/after FcɛRI-stimulation. IKK2 and SNAP- 23 co-immunoprecipitated in the raft fractions following FcɛRI-stimulation (Fig. 8). These results suggest that IKK2 associates with SNAP-23 in the lipid rafts only after FcɛRI-stimulation.
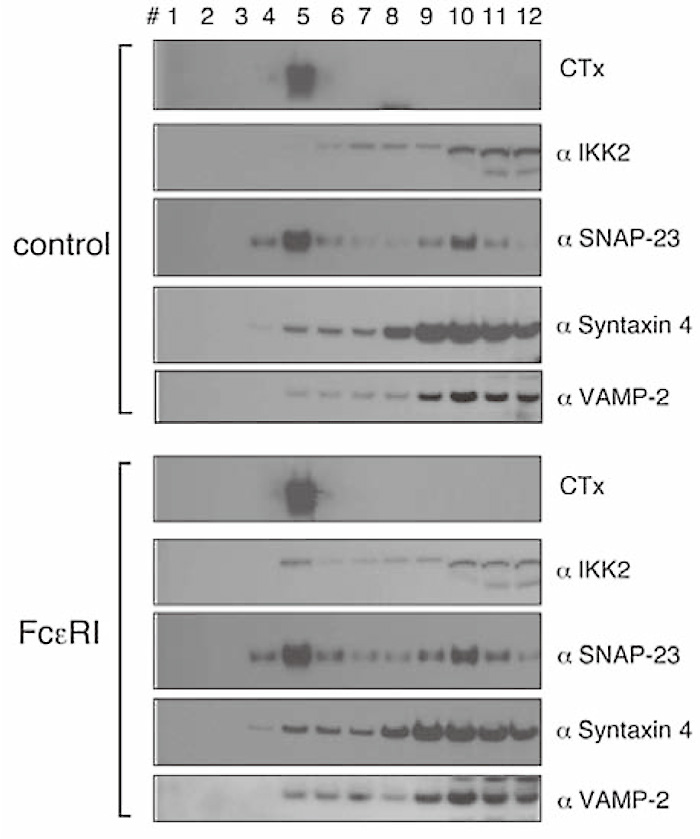
Fig. 7
IKK2 is recruited to the lipid rafts upon FcɛRI stimulation.
Localization of IKK2, SNAP-23, Syntaxin 4, and VAMP-2 in lipid raft upon FceRI stimulation. Cell lysates of unstimulated or stimulated RBL-2H3 cells were fractionated by sucrose density gradient ultracentrifugation and blotted with each Ab. CTx blotting indicates that fraction #5 contains lipid raft.( ref.3)
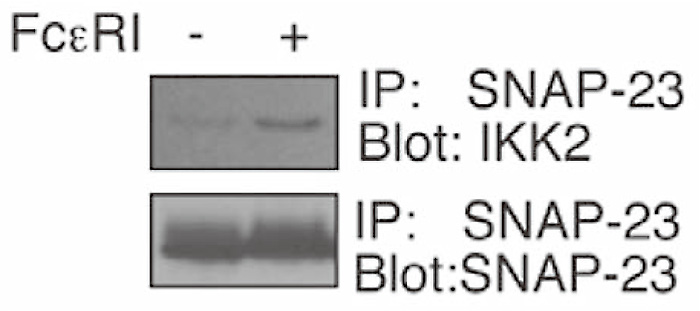
Fig. 8
IKK2 associates with SNAP-23 upon FcɛRI stimulation.
IKK2 associates with SNAP-23 in the lipid raft fractions only after FcɛRI-stimulation. Lipid raft fraction isolated from unstimulated or stimulated RBL-2H3 cells were subjected to immunoprecipitations (IP) with anti-SNAP-23 antibody. Immunoprecipitants were blotted with each Ab. (ref.3)
Phosphorylation of SNAP-23 on Ser120 and Ser95 modulates regulated exocytosis by mast cells[2]. I hypothesized that SNAP-23 is a substrate for IKK2. In vitro kinase assays showed that IKK2 phosphorylated SNAP-23 (Fig. 9A). However, when Ser120, Ser95, or both of them were converted to Ala, phosphorylation of recombinant mutant SNAP-23 by IKK2 was substantially attenuated (Fig. 9A), indicating that IKK2 can phosphorylate SNAP-23 at both Ser120 and Ser95 in vitro. To assess whether this phosphorylation by IKK2 occurs in vivo, we used phospho-specific antibodies.
Ser 120 and Ser95 were phosophorylated upon FcɛRI stimulation in wild type (Fig. 9B). In contrast, little or no phosphorylation was detected in IKK2-/- FLMCs (Fig. 9B). These results show that SNAP-23 is phosphorylated by IKK2 at Ser120 and Ser95 in vitro and in vivo and that IgE-induced phosphorylation of SNAP-23 requires IKK2.
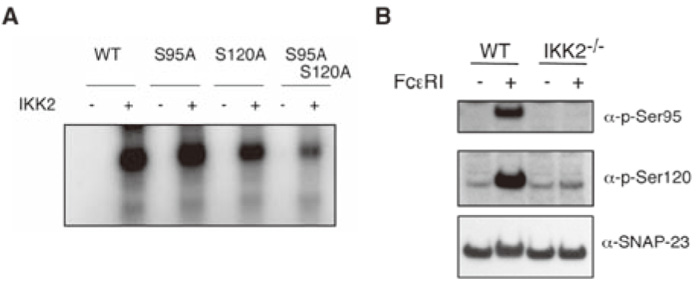
Fig. 9
IKK2 phosphorylates SNAP-23 on Ser120 and Ser95.
(A) Equal amounts of purified wild type and mutant GST-SNAP-23 proteins were incubated with[ γ-32P] ATP and IKK2 and the reaction was analyzed by SDSPAGE and PhosphorImager analysis.( B) IgE-induced phosphorylation of SNAP-23 on Ser120 and Ser95 is impaired in IKK2-/- mast cells. WT or IKK2-/- mast cells were cross-linked upon FceRI. Fifteen minutes later, cell lysates were recovered and analyzed by immunoblot analysis using phosphorylation site specific antibodies. (ref.3)
Finally, to examine the effect of expression of phospho-mimetic mutant of SNAP-23, which can function without serine phosphorylation, in IKK2-/- mast cells on FcɛRI-stimulated β-hexosaminidase release and in vivo PCA reactions, phospho-mimetic mutant SNAP23EE by mutating serines 95 and 120 was generated. Retroviral transduction of WT SNAP- 23 did not rescue the impairment of FcɛRI-induced β-hexosaminidase release in IKK2-/- mast cells (Fig. 10A). By contrast, expression of SNAP-23EE partially rescued the impairment of FcɛRI-induced degranulation in IKK2-/- mast cells (Fig. 10A).
I also examined in vivo early phase PCA reactions in mast cell-reconstituted W/Wv mice. Mice with WT SNAP- 23 or mock transduced IKK2-/- FLMCs exhibited similar levels of reduced cutaneous anaphylaxis (Fig. 10B), in comparison with mice with control WT FLMCs, as assessed by change in ear thickness 1 hour after antigen challenge. By contrast, mice with SNAP- 23EE transduced IKK2-/- FLMCs exhibited increased cutaneous anaphylaxis than mice with mock transduced IKK2-/- FLMCs (Fig. 10B). These results suggest that lack of phosphorylation of SNAP-23 on Ser120 and ser95 is responsible for the impairment of FcɛsRIinduced degranulation in IKK2-/- mast cells. Taken together, these results indicate that IKK2-induced SNAP-23 phosphorylation is a key event in SNARE complex formation leading to IgE-induced degranulation in mast cells (Fig. 11).
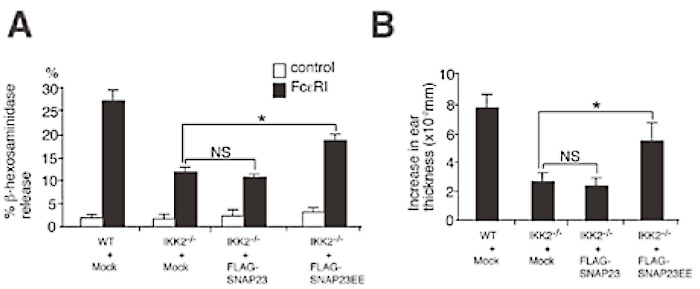
Fig. 10
Phosphorylation of SNAP-23 on Ser120 and Ser95 involved in IKK2-mediateddegranulation.
(A) Expression of phospho-mimetic mutant of SNAP-23 partially rescue the impairment of FceRI-induced β-hexosaminidase release in IKK2-/- mast cells. Mast cells were infected with retroviruses of pMX-puro-FLAGWT SANP-23, pMX-puro-FLAG-SNAP-23EE, or pMXpuro (Mock). Cells were cross-linked upon FcɛRI.
Thirty minutes later, β-hexosaminidase activity in the supernatants and lysates of the cell pelletes was quantified.*P<0.01. NS, not significant. (B) For passive cutaneous anaphylaxis, mock vector transduced WT mast cells, and mock vector, WT SNAP-23, or SNAP-23EE transduced IKK2-/- mast cells, were injected intradermally into the right ear of W/Wv mice. Four weeks later, PCA were induced in these mice. *P<0.01. NS, not significant. (ref.3)
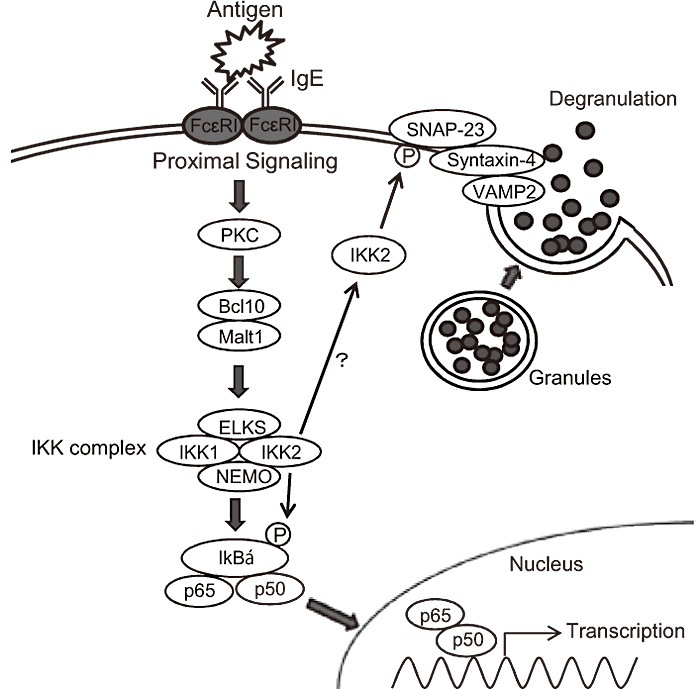
Fig. 11
IKK2 plays critical roles in mast cell degranulation.
IKK2-induced SNAP-23 phosphorylation is a key event in SNARE complex formation leading to IgE-induced degranulation in mast cells.
In addition to critical roles of IKK2 in NF-κBmediated biological responses, my data has identified novel NF-κB-independent roles of IKK2 in mast cell degranulation. A thorough characterization of IKK2 in mast cell degranulation will provide new insights into the pathogenesis of mast cell-mediated allergic diseases such as asthma, allergic rhinitis, and atopic dermatitis. I have already demonstrated that IKK2-selective inhibitors repress mast cell degranulation, suggesting that IKK2- selective inhibitors have a therapeutic potential for allergic diseases.
I thank Dr. I. M. Verma and Dr. H. Nakajima for valuable discussion, useful comments and helpful supports.
Address correspondence to Dr. Kotaro Suzuki.
Department of Allergy and Clinical Immunology, Graduate School of Medicine, Chiba University, 1-8-1 Inohana Chuouku, Chiba 260-8670, Japan.
Phone: +81-43-226-2198. Fax: +81-43-226-2199.
E-mail
: suzuki_k@faculty.chiba-u.jp
