




Volume 84, Number 2
doi:10.20776/S03035476-84-2-P75
[Original Paper]
Zhongqiu Ji, Shigeru Sugaya and Nobuo Suzuki
(Received December 3, 2007, Accepted December 12, 2007)
It is important to search for mechanisms which suppress the mutagenicity of environmental agents in human cells and to utilize cultured human cells which are suitable for this search. In this study a human RSa cell line was used for the detection of cadmium mutagenicity. RSa cells were already found to be hypermutable after treatment with various agents, as assessed by both phenotypic and genetic mutation tests, ouabain-resistance (OuaR) tests and differential dot-blot hybridization tests.
Treatment of RSa cells with cadmium resulted in a 1.97 fold increase in the frequency of OuaR mutation in comparison with the frequency with mock treatment. In contrast, UVr-1 cells derived from RSa cells as a UV-resistant variant cells, showed slightly increase of OuaR mutation frequency after cadmium treatment. Moreover, K-ras codon 12 mutations in genomic DNA from cadmium-treated RSa cells were detected by polymerase chain reaction following differential dot blot hybridization, whereas the mutation was not detected in UVr-1 cells. DNA damage, which was detected by comet assay, was detected after cadmium treatment in both RSa and UVr-1 cells. There is no difference of excision repair capacity between RSa and UVr-1 cells. Thus, RSa cells are hypermutable for cadmium and DNA repair but for excision repair may be related with the hypermutability.
Cadmium, Human cells, Mutation, K-ras, Comet assay
Poisonous materials containing heavy metals, such as cadmium deposited in the earth, will cause various pollutions. Cadmium can cause osteomalacia, bone distortion, bone fracture and even cancer. Subsequently, it is essential for us to develop a method that will conveniently evaluate the unhealthy effects of such poisonous materials, including the mutagenic activity of cadmium on human cells [1].
Many researchers have tried to establish a method of identifying the biological effects of cadmium using mammalian cells. For example, Oxidative DNA damage, 8-OhdG, and mutation are positively associated with cadmium exposure in mice [2]. However, no reports can be found concerning the molecular mechanisms of cadmium mutagenicity upon human cells [3]. Thus, it is necessary to develop a method which is both applicable and useful in giving us a comprehensive understanding of the mutagenic effects of cadmium.
In the present study, the mutagenic effects of cadmium on human RSa cells were estimated by using ouabain-resistant phenotypic mutation assay and by K-ras codon 12 base substitution mutation assay. The latter assay was assessed by PCR and differential dot-blot hybridization [4]. A human embryo-derived RSa cell line has been utilized for the investigation of mutagenic agents because of its hypermutable potential [5]. The hyper mutability of RSa cells is due possibly to the low capacity of repair synthesis for damaged DNA, so-called excision repair [6]. In fact, human interferon induces an increase of DNA repair capacity in RSa cells, in association with the suppression of mutation [7]. It is significant for us to find out the mutagenicity of human RSa using only a few amounts of environmental agents with DNA-damaging activity. So in this case it was determined whether or not cadmium treatment causes DNA damage in RSa cells.
When the biological effects of environmental agents are investigated, not only chemical compounds but also UVC (ultraviolet rays with principally 254 nm wavelength) produces valuable information. In addition to excision repair, other DNA repair mechanisms are suggested by studying the cellular responses to chemical and/or UVC treatment [8]. For investigation of the UVC responses in human cells UVr-1 cells were previously established from RSa cells as a variant with an increased resistance to UVC cell-killing and without any differences regarding excision repair capacity [9]. A comparison of the responses between UVr-1 cells and the parent RSa cells after treatment with DNA damaging agents have resulted in new findings concerning the cellular molecules involved in the responses [10-14]. In order to determine the impact of cadmium on the mutation ability of cells and to establish a practical method of monitoring the cadmium influence on human cells, UVr-1 cell were also utilized in detecting mutagenicity and searching for the suppression of mutagenicity.
Cadmium chloride and other agents were purchased from Wako Pure Chemical Industry (Osaka, Japan).
Human RSa cells have been characterized as having a high susceptibility to mutagens [15]. UVr-1 cells have been described elsewhere [8]. These cells were cultured in the following medium; EMEM (GIBCO/BRL, Grand Island, NY) containing 5% (v/v) calf serum (CS; Intergen, Purchase, NY) , at 37℃ in a humidified atmosphere containing 5% (v/v) CO2.
After a 24 h inoculation of the cells (3×105 cells per one 60mm dish) , they were then cultured in the medium containing cadmium for 24 hours. Twenty four hours after the cadmium treatment, the cells were re-seeded for mutation assay, as described elsewhere [7].
Cells in the logarithmic growth phase were treated with and without cadmium. Cells were seeded in each well of 96-well plates (5×103 cells/well) and cultured for 24 h. The activity of mitochondrial succinic dehydrogenases were measured by incubating the cells for 4 h in the presence of MTT [3- (4, 5-dimethylthiazol-2-yl) -2, 5-diphenyl tetrazolium bromide] (0.5 mg/ml) followed by a measurement of absorbance at 570 nm with a reference wavelength of 655 nm according to the method of Mosmann [16], as described previously [10]. The absorbance, reflecting the viable cell number after cadmium treatment, was expressed as a percentage of the entire test subject group, including those cells which were mock- treated (without cadmium exposure).
The survival capacity of RSa and UVr-1 cells treated with and without cadmium was measured by a colony formation assay principally accorded to the method described previously [17]. In summation, cells were inoculated at a density of 1×103 cells per 100mm dish and 24 h after the inoculation cultured in MEM with and without cadmium. After the cadmium treatment or mock treatment, cells were cultured with the aforementioned medium for 14 days followed by fixation and staining with a 30% methanol-water solution containing 0.2% (w/v) methylene blue. Colonies containing approximately 50 or more cells were counted and then survival percents were estimated as a relative ratio to those which had undergone mock treatment.
Base substitution mutation of codon 12 of K-ras was performed according to the PCR and differential dot-blot hybridization methods, as described previously [18]. In summation, the cells were inoculated with and without cadmium for 24 hours. Then 6 days after genomic DNA was extracted from the cells using a standard proteinase K/SDS/phenol chloroform procedure. Genomic DNA from the cells was amplified by PCR using the primers for the codon 12 of K-ras, 5’-ACTGAATATAAACTTGTGG-3’and 3’-GCTTATACTAGGTTGTTATC-5’. The amplified products were dot-blotted onto nylon membranes. After hybridization with digoxigenin-11-dUTP-3’-end-labeled K-ras codon 12 probes, the membranes were washed and blocked with a blocking reagent. They were then incubated with polyclonal sheep anti-Dig Fab conjugated to alkaline phosphate (Boehringer Mannheim, Germany) and colored with nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate solution (Boehringer Ingelheim, Germany). As a normal probe, the oligonucleotide 5’-GTTGGAGCTGGTGGCGTAGG-3’ was used. However, mutant probes contained the following oligonucleotides which were mixed at the same concentration ratios: 5’-GTTGGA-GCTAGTGGCGTAGG-3’, 5’-GTTGGAGCTCG-TGGCGTAGG-3’, 5’-GTTGGAGCTTGTGGCG-TAGG-3’, 5’-GTTGGAGCTGATGGCGTAGG-3’, 5’-GTTGGAGCTGCTGGCGTAGG-3’ and 5’- GTTGGAGCTGTTGGCGTAGG-3’.
Quantitative assay for the induced frequency of OuaR cells was carried out following the methods described previously [7]. The cells underwent cadmium treatment for 24 hours were plated at a density of 8×104 cells per one 100mm dish in a medium containing 6×10-8 M ouabain. Every second day the medium was changed and 21 days later the colonies were counted. For the estimation of cloning efficiency, 8×102 cells per one 100mm dish were also cultured without ouabain, and 14 days later the colonies were counted. The mutation frequency was determined by dividing the total number of OuaR mutant colonies by the total number of cells plated, corrected by the cloning efficiency and expressed as mutants per 105cells.
The comet assay was performed according to a method of single cell gel electrophoresis assay (Trevigen, Gaithersburg, MD) [19]. In summation, treatment of the cells were done with and without cadmium for 24 hours, followed by lyzation of cells using lyses solution. After the lyzation, the slides were placed on a horizontal gel electrophoresis unit filled with fresh electrophoresis buffer (300mM NaOH, 1mM Na2EDTA, PH>13). They were then left for 60 min for DNA unwinding and then subjected to electrophoresis for 40 min at 1.25V/cm (300mA). The electrophoresis was performed under dim light. After electrophoresis, the slides were washed with a neutralizing buffer (0.4 M Tris, pH7.5) and the cells were stained with SYBR Green I (Molecular Probes, Eugene, OR, USA). Observations were made with a magnification using a fluorescent microscope (Olympus equipped with a 515-560nm excitation filter) connected through color video camera. The image for each individual cell was acquired immediately after opening the microscope shutter to the computer monitor, following the evaluation of comet moment using Scion image software (Scion co. Frederick, MD). Tail moment and tail intensity were used to determine the DNA damage. Tail moment, defined by the product of the distance between the head and the tail and the proportion of DNA in the tail, was used to evaluate the extent of DNA migration. The rate of comet moment was obtained from number six cells at each dose of cadmium.
Sensitivity to cadmium-induced cell death was first examined by MTT and colony survival assay in RSa and UVr-1 cells. RSa cells had higher sensitivity to cadmium-induced cell death than did UVr-1 cell (Fig. 1).
Using RSa cells, the mutagenic effect of cadmium was tested by phenotypic OuaR mutation assay. As shown in Fig. 2, the survivors of OuaR colonies increased in cells treated with 10-7 M and 10-6 M cadmium for 24 hours. Using the same concentrations as examined in OuaR mutation assay, base substitution mutations at K-ras codon 12 in genomic DNA from RSa cells exposed to cadmium were analyzed, and hybridization signals with K-ras codon 12 mutant probes were detected (Fig. 3).
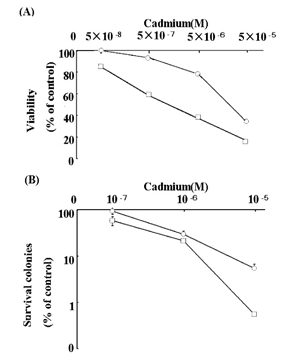
Fig. 1
Cell viability (A) and colony survival (B) was evaluated after treatment with and without cadmium in RSa cells (□) and UVr-1 cells (Ο). Cells were treated with and without cadmium and viability and colony survival were estimated as described in Materials and Methods. The data represents the average percentage of numbers relative to those of mock-treated cells, and are the means ± S. E. M. of three independent experiments.
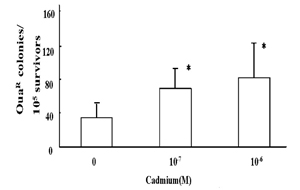
Fig. 2
OuaR mutation frequency in RSa cells treated with and without cadmium for 24 hours. Quantitative assay for the induced frequency of OuaR cells was carried out as described in Materials and Method. *, P<0.05, the cadmium treatment vs. the mock treatment.

Fig. 3
K-ras codon 12 mutation by PCR and differential dot-blot hybridization of RSa cells with and without cadmium treatment. The differential dot-blot hybridization was performed after treatment of RSa cells with cadmium, as described in Materials and Methods.
Cadmium mutagenicity was furthermore examined in a RSa cell-derived variant, UVr-1, under the same assay conditions as examined in the parent RSa cells. UVr-1 cells showed only a slight increase of OuaR colonies at 10-7 and 10-6 M cadmium (Fig. 4A). On the other hand, hybridization signals with K-ras codon 12 mutant probes were not detected in the UVr-1cells treated with the agent (Fig. 4B).
In cadmium-treated RSa and UVr-1 cells, cellular DNA damage was examined using comet assay. Comet moment increased in RSa cells treated with cadmium for 24 hours at 10-7 and 10-6 M, whereas a small increase was detected in UVr-1 cells at the same condition of cadmium treatment (Fig. 5). Thus, DNA damage was detectable in both cells after cadmium treatment.
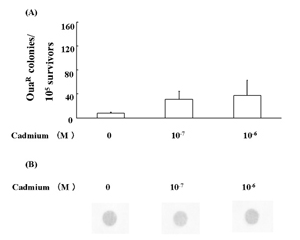
Fig. 4
Low susceptibility of UVr-1 cells to cadmium mutagenicity. Quantitative assay for the induced frequency of OuaR cells (A) and K-ras codon 12 mutation analysis (B) were performed as described in Materials and Methods. *, P<0.05, the cadmium treatment vs. the mock treatment.
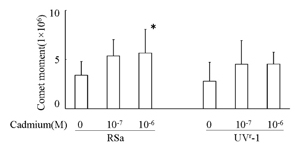
Fig. 5
Effects of cadmium on DNA damage using comet assay in RSa and UVr-1 cells. Comet assay was performed as described in Materials and Methods. *, P<0.05 the cadmium treatment vs. the mock treatment.
The capacity of cell survival after cadmium treatment was found to differ between the RSa and variant UVr-1 cells (Fig. 1). RSa cells appear to have a low capacity repair function for damaged DNA [6]. The single cell gel electrophoresis, named as the comet assay, has been introduced as a simple method to measure DNA damage. This comet analysis revealed that cadmium treatment results in cellular DNA damage in RSa cells (Fig. 2). Due to DNA damage RSa cells with low repair-capacity seemed to have low levels of cell survival after cadmium treatment (Fig. 1).
The deficient DNA repair mechanism in RSa cells is thought to be excision repair [6]. The variant UVr-1 cells also have the same low capacity of DNA repair and possibly excision repair, even though those cells are more resistant to UVC cell-killing effects than the parent RSa cells [9]. Interestingly, UVr-1 cells have a more drastic recovery of DNA synthesis from UVC-caused suppression, and therefore UVr-1 cells may have more activity than other DNA repair mechanisms except for excision repair of RSa cells [8,9]. The unknown DNA repair may have contributed to recovery of UVr-1 cells from cadmium-induced DNA damage and subsequent cell-killing. Thus, it seems likely that UVr-1 cells are more resistant to cadmium cell-killing than RSa cells (Fig. 1), and that comet moment in cadmium-treated UVr-1 cells has decreased in a greater extent than RSa cells (Fig. 5).
In the present study, it has been demonstrated that cadmium treatment results in the induction of OuaR and K-ras codon 12 mutations in RSa cells (Fig. 2 and 3). To determine the sensitivity of the K-ras codon 12 mutation assay, the mutation of genomic DNA from SW480 cells carrying K-ras codon 12 mutations was previously compared by the dilution of the DNA into human placental DNA, followed by amplification of the target sequences [20]. Oligonucleotides amplified both from SW480 cells and human placental DNA were hybridized using only mutant probes and normal probes, respectively. The mutant sequences of the antisense strand of K-ras codon 12 which were expected to be detected by the mutant probe used, were ACT, ACG, ACA, ATC, AGC and AAC. The amplified oligonucleotides from the DNA preparation containing more than 0.1% genomic DNA from SW480 cells were inferred by a hybridization signal with the mutant probes. Hybridization signals with K-ras codon 12 mutant probes were detected in RSa cells treated with cadmium at 10-7 M and 10-6 M (Fig. 3). Therefore, more than one RSa cell per 103 cells may have been susceptible to cadmium-induced K-ras codon 12 mutation events. Also, according to the frequency of OuaR mutation analysis (Fig. 2) more than 7 cells per every 104 survival cells may have been detectable as OuaR mutation cells.
In contrast, UVr-1 cells showed less than 3 OuaR mutated cells per 104 survival cells (Fig. 4A) and undetectable K-ras codon 12 mutations (Fig. 4B). These different levels of induced mutation frequency between OuaR and K-ras analysis might be due to a difference of targeted genes, as phenotypic OuaR mutation occurs from Na+ and K+-ATPase gene mutations [18]. On the other hand, the low or undetectable mutations in UVr-1 cells may be due to some repair mechanisms with the exception of excision repair. The unknown repair mechanisms may also have contributed to only slightly increased levels of comet moment in UVr-1 cells (Fig. 5).
The involvement of HSP27 regarding a resistance to UVC lethality was recently found in comparative studies using UVC-sensitive human RSa cells and a variant of UVC-resistant variant APr-1 cells, followed by an analysis using samples taken from cells containing sense and antisense cDNA for HSP27 [21]. Wano et al. reported on the genes responsible for the sensitivity to UVC-induced cell death [21]. Therefore, we can conduct a preliminary examination regarding the comparison of gene expression levels using cDNA array analysis between RSa and UVr-1 cells. Among 22000 cDNAs examined, 112 genes were up-regulated more than four fold in UVr-1 cells, although 437 genes were down-regulated in less than three fold of UVr-1 cells compared with RSa cells. Based on the results of genes with discrepancies in their expression levels between the two cell lines, including HSP27 expression levels, the repair mechanisms underlying inconsistencies regarding cadmium susceptibility between RSa and UVr-1 cells need to be investigated for further clarification. Along with advancements in techniques based on cell analysis, a highly sensitive method, which clarifies mutagenicity of environmental agents in human RSa cells and the variant UVr-1 cells, is now expected to open up the way for new research disciplines such as human cell molecular biology.
This work was supported in part by grants-in aid from the following foundations: the Smoking Research Foundation, the Tokyu Foundation for Better Environment, “Ground Research for Space Utilization” promoted by NASDA and the Japan Space Forum and the Ministry of Education, Science and Culture (Japan).
【目的】
化合物によるヒト細胞での変異原活性を高感度で検出し,その変異誘導メカニズムを探究できる方法の開発が希求されている。ヒト細胞へ致死毒性や他生物での変異原活性などの報告があるカドミウムについては,ヒト細胞における変異原活性とそのメカニズムに関する詳細な報告が未だない。そこで,変異原活性の検出に有用な培養ヒト細胞を利用することにより,カドミウムの変異原活性を高感度で検出可能とさせ,その活性の特異な点を明らかにすることとした。
【方法】
培養ヒト細胞RSaとその派生株UVr-1を使用した。変異原活性の検出には,ウワバイン耐性化形質変異の検出法と癌遺伝子K-rasのコドン12における塩基置換変異を検出する方法を併用した。遺伝子傷害については,コメットアッセイ法を用いて解析した。
【結果】
RSa細胞におけるカドミウムの変異原活性はウワバイン耐性化と塩基置換の両変異で検出され,他方,UVr-1細胞ではウワバイン耐性化でわずかな変異誘導がみられたが塩基置換変量は検出されなかった。但し,両細胞と共にDNA傷害が誘導された。
【結語】
培養ヒト細胞RSaを用い,DNA傷害を介してのカドミウムの変異原活性を用い,高感度で検出可能とすることが示唆された。RSa細胞とUVr-1細胞では除去修復のレベルに相違がないことから,カドミウムの変異原活性を抑える除去修復以外の他のメカニズムがあるとの示唆が得られた。
Department of Environmental Biochemistry, Graduate School of Medicine, Chiba University, Chiba 260-8670.
紀 仲秋,菅谷 茂,鈴木信夫: ヒト細胞RSaと派生株UVr-1におけるカドミウム変異誘導に対する感受性の相違.
千葉大学大学院医学研究院環境影響生化学
Tel & Fax. 043-226-2041. E-mail: nobuo@faculty.chiba-u.jp
2007年12月3日受付,2007年12月12日受理.SUMMARY
