




Volume 84, Number 4
doi:10.20776/S03035476-84-4-P179
[Original Paper]
Kenshi Kawasaki, Katsuhiro Uzawa, Yoshinori Kurasawa, Naruhide Yoshida
Ken Shimada, Hisako Uesugi, Akiyuki Murano, Yukio Hayashi, Makoto Yamaki
Tetsuhiro Moriya, Masashi Shiiba and Hideki Tanzawa
(Received February 27, 2008, Accepted April 2, 2008)
Headpin, which is located at chromosome 18q21.3, belongs to serine proteinase inhibitor family (serpin). One of the functions of this gene family is believed to inhibit hemangiogenesis, invasion, and metastasis of tumors. In this study, 30 clinical tissue specimens of oral squamous cell carcinoma (OSCC) and 9 OSCC-derived cell lines were examined for the down regulation of the gene expression. In addition, the restoration of headpin mRNA expression induced by 5-Aza-2'-deoxycytidine (5-AzaC) treatment, a DNA demethylating agent, was evaluated in OSCC-derived cell lines. RT-PCR revealed the down-regulation of headpin gene expression in 17 (56.7%) of 30 primary OSCC samples and in 5 (55.6%) ; SAS, HSC-3, OK92, HO-1-N-1 and SCC4, of 9 OSCC-derived cell lines. Among those 5 cell lines, 5-AzaC treatment induced the restoration of headpin gene expression in 4 (80.0%) cell lines; SAS, HSC-3, OK92 and SCC4. These results suggested that the down-regulation of headpin expression contributes to the development of human OSCC and that DNA hypermethylation is one of the important mechanisms inactivating the headpin gene in oral carcinogenesis.
oral squamous cell carcinoma (OSCC), headpin gene, serpin family, methylation, demethylation, 5-Aza-2'-deoxycytidine (5-AzaC)
Serine proteinase inhibitors (serpin) consist of structurally related proteins participating in the regulation of complex physiological processes and contain over 500 members in the genomes of most metazoan, plants, and certain viruses [1,2]. In humans, more than 35 serpin members are known to participate in extracellular and intracellular physiological processes. The primary function of serpin family members appears to be neutralizing overexpressed serine proteinase activity and serpins play an important role in blood coagulation, inflammation, cell migration, fibrinolysis, complement activation, remodeling of extracellular matrix, hormone transport, hemangiogenesis, apoptosis, cancer growth, invasion, and metastasis [1,2,3]. A novel serpin named headpin was recently identified, cloned and mapped to chromosome 18q21.3 [4,5]. The cloning of hurpin/P113, which has a sequence identical to that of headpin, was also reported [6]. Serpin families tend to cluster at specific chromosomal locations. The ov-serpin family, including headpin, maspin, SCCA1, SCCA2, and PAI2, clusters at 18q21 [7,8] and several reports, including our previous paper, indicated frequent loss of heterozygosity at the 18q21-23 chromosomal region in head and neck carcinomas and other solid malignancies [9-12]. There have been only few reports validating the roles of headpin gene in many clinical samples and tumor cell lines, especially in OSCC.
In the present study, in order to clarify the role of the headpin gene in OSCC, 30 clinical tissue specimens of OSCC and 9 OSCC-derived cell lines were examined for the status of the gene expression level. Moreover, the restoration of headpin mRNA expression by 5-AzaC treatment was evaluated in OSCC-derived cell lines.
Clinical tissue samples.
Thirty pairs of primary OSCC tissue samples and adjacent normal oral epithelia were obtained at the time of surgical resection from 1995 to 2001 at Chiba University Hospital. Informed consent was obtained from all patients and the patient's families. One part of each tissue sample was frozen and stored at -80℃ for RNA extraction, and the remaining tissues were examined microscopically to confirm the diagnosis and to evaluate tissue morphology. The histopathological classification was performed according to the International Histological Classification of Tumors and the clinical staging was determined by the UICC TNM staging system. The patients, from whom the tissue specimens were obtained, were well informed and consented to offer their tissues for this study.
Cell lines
The following OSCC-derived cell lines were analyzed: SAS, HSC-2, HSC-3, HSC-4, Ca9-22, HO-1-u-1, HO-1-N-1, SCC4 (Human Science Research Resource Bank, Osaka, Japan) and OK92 (established in our department from a carcinoma of the tongue). All cell lines were cultured in RPMI-1640 medium with 10% fetal bovine serum and 50 units/ml penicillin and streptomycin in an atmosphere of 5% CO2 at 37℃.
RNA extraction
Total RNA was extracted from cell lines and tissue samples using Trizol (Invitrogen Life Technologies, CA USA) according to the manufacture's instructions. Extracted RNAs were subjected to Affimetrix GeneChip hybridization or cDNA synthesis.
Affimetrix GeneChip hybridization
Double-stranded cDNA was synthesized from 20 μg of total RNA using the Superscript Choice system (Invitrogen). After phenol/chloroform extraction and ethanol precipitation, a biotin-labeled in vitro transcription reaction was carried out using the cDNA template (Enzo Bioarray, Farmingdale, NY, USA). Seven microgram of cRNA was fragmented according to Affimetrix protocols and added to the recommended hybridization mixture. Expression profiles were created using the Human Genome U 133 Plus 2.0 arrays containing 54,675 probe sets (Affimetrix, Santa Clara, CA, USA). Arrays were stained with phycoerythrin-streptavidin antibody followed by a second staining with phycoerythrin-streptavidin. Arrays stained a second time were scanned using the Affimetrix GeneChip Operating Software 1.1 (Affimetrix) and then genes of the serpin superfamily were classified by GeneSpring 6.1 (Silicon Genetics, Redwood City, CA, USA).
Evaluation of suppressed expression of headpin gene by semiquantitative RT-PCR method.
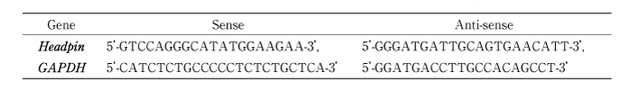
PCR reactions were performed in a 9700 Perkin-Elmer Thermal Cycler at 94℃ for 1 min, with 35cycles of 94℃ for 1 min, 55℃ for 90s, and 72℃ for 90s, followed by an extension step at 72℃ for 10 min. After amplification, an aliquot of the PCR product was separated on a 3% TAE-agarose gel and staind with ethidium bromide. All subsequent assays were carried out using parameters that yielded amplification of both the headpin and GAPDH genes within a linear range. cDNA was amplified by PCR using primers specific for cDNA of the headpin gene (Table 1). And the GAPDH gene was also amplified using specific primers as an internal control (Table 1) [13]. The density of the ethidium bromide-stained bands was analyzed using the NIH Image software. The results were normalized as a ratio of each specific mRNA signal to the GAPDH gene signal within the same RNA sample. cDNA obtained from normal oral epithelium was used as a positive control. Reproducibility was confirmed by processing all samples at least twice.
Evaluation of headpin mRNA expression in OSCC-derived cell lines by 5-Aza-2'-deoxycytidine treatment.
We examined the promoter region of headpin gene for CpG island. However, no CpG island was found in the region of 2100 residues upstream from exon 1. Therefore we're not able to perform methylation specific RT-PCR method.
To assess restoration of headpin expression, the cells were treated with 2μM of the 5-AzaC as described previously [14]. On the day 5, the cells were washed with PBS and grown for another 10 days without 5-AzaC. After total RNA isolation from these samples, RT-PCR analysis was performed with the same procedures as described above.
Table 1
Sequences of specific primers used in RT-PCR analyses.
Microarray analysis.
To detect down-regulated genes in OSCC, microarray analysis was performed using 4 OSCC samples. The data showed that there were 13 genes which were down-regulated in all of 4 samples. Table 2 summarizes the list of those genes (Table 2). Among them, we focused on headpin gene and we analyzed the association of this gene in OSCC.
The status of headpin gene in primary OSCC tissue specimens and OSCC-derived cell lines.
Semi-quantitative RT-PCR of headpin gene was performed. We examined 30 cases of oral SCC and 9 cell lines derived from oral SCC. mRNA expression of headpin was not observed in 4 of 9 cell lines; SAS, HSC-3, HO-1-N-1, SCC4, and slight expression was detected in OK92 cell line (Fig. 1). And 17 (56.7%) of 30 cases showed much suppressed expression of headpin gene (Fig. 2). However, significant correlation between clinicopathological characteristics and headpin expression was not indicated (Table 3).
Evaluation of headpin mRNA expression in OSCC-derived cell lines by 5-AzaC treatment.
In order to determine whether methylation might inhibit headpin mRNA expression, 5 cell lines in which showed headpin expression was down-regulated; SAS, HSC-3, OK92, HO-1-N-1 and SCC4, were exposed to demethylating agent 5-AzaC, which could restore mRNA expression suppressed by gene methylation. After treatment with 5-AzaC, the expression of headpin mRNA was restored in SAS, HSC-3, OK92 and SCC4 (Fig. 3). The data suggested that hyper-methylation was closely related with down-regulated expression of headpin gene in OSCC.
Table 2
Down-regulated genes in OSCC samples detected by microarray analysis.

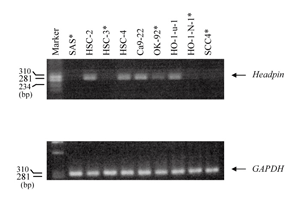
Fig. 1
mRNA expression of headpin gene in OSCC-derived cell lines analyzed by RT-PCR.
Headpin mRNA expression was not observed in 4 cell lines (SAS, HSC-3, HO-1-N-1, SCC4) and slight expression was shown in OK92 cell line. Asterisks show cell lines in which headpin mRNA expression was down-regulated.
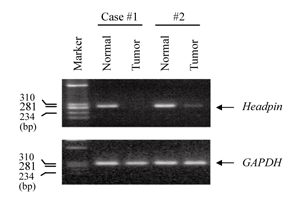
Fig. 2
Representative results of RT-PCR analysis for the down-regulated expression of headpin gene in clinical OSCC tissue specimens.
mRNA expressions of headpin gene were significantly down-regulated in tumor samples compared with normal samples.
Table 3
Correlation between clinicopathological characteristics and down-regulation of headpin gene.
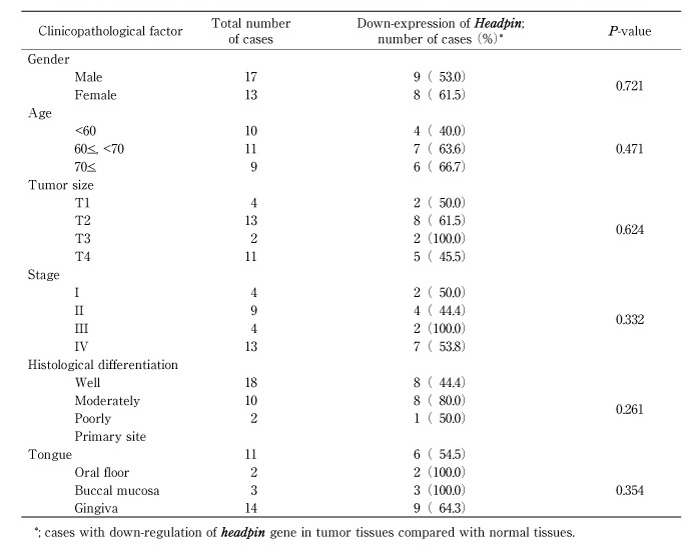
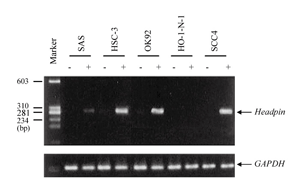
Fig. 3
RT-PCR analysis for the demethylation study of OSCC-derived cell lines. After treatment with 5-AzaC, restoration of headpin mRNA expression was found in the 4 cell lines; SAS, HSC-3, OK-92 and SCC4, of 5 ones which showed down-regulation of the gene before the treatment. "-", before 5-AzaC treatment; "+", after 5-AzaC treatment; GAPDH, internal marker.
cDNA microarray identified 13 genes with down regulated mRNA expression in OSCC, comparing with normal oral epithelium. Out of these genes, headpin gene was singled out as a candidate gene for a tumor suppressor gene associated with oral SCC, because of its molecular function; heamangiogenesis, invasion and metastasis (Table 2). The high frequency of down regulated expression of headpin gene in primary oral SCC specimens supported this hypothesis and suggested the important role of the gene in oncogenesis of oral SCC. However, no significant relationship between the down expression of the gene and the clinicopathological features was found in our results. Thus, further investigations should be necessary to clarify clinical roles of down-expression of headpin gene.
Headpin is a novel serine proteinase inhibitor and localized at chromosome 18q21.3, which is expressed in normal squamous epithelium of the oral mucosa, skin and cervix [4]. Inhibitory serpins are known to play an important role in tumor invasion, metastasis, tumor suppression and apoptosis [5-7], and their sequences have homology with the ovalbumin serpins (ov-serpins) which include the squamous cell carcinoma antigens (SCCA1, SCCA2), plasminogen activator inhibitor-2 (PAI2) and maspin. In humans, the ov-serpins share 40% amino acid sequence identity, contain seven or eight exons, lack typical signal peptides and map to one of two chromosomal clusters. P16, p19 and ELNAH2 map to 6p25, whereas p15 (maspin), SCCA2, SCCA1, PAI2, PI10 and p18 map to within a 500kb interval of 18q21.3 [8]. Deletion and LOH at 18q21 are frequent in several types of tumors [9,10]. Moreover, our data suggest that hypermethylation of headpin is one of the crucial mechanisms inducing the down-regulation of headpin gene expression. These results indicate that the clustering of ov-serpins at 18q21.3 and hypermethylation assay may have important diagnostic considerations when assessing tumor phenotypes. In this study, there was no relationship between the status of the gene expression and clinicopathological findings. Therefore, further study will be necessary to establish the diagnostic method of assessing tumor phenotypes.
We thank Lynda C. Charters and Allan Earle for proofreading this manuscript.
Headpinは染色体18q21.3上に位置するセリン・プロテアーゼ・インヒビター (serpin) ファミリーの一遺伝子である。この遺伝子ファミリーは腫瘍における血管新生,浸潤,転移の抑制を行うとされている。本研究では30の口腔扁平上皮癌臨床組織検体と9種の口腔癌由来細胞株におけるheadpinの発現について調べた。さらに脱メチル化剤5-AzaC処理による発現減弱の回復について検討した。RT-PCR解析では30の臨床組織検体のうち17検体 (56.7%) ,9種の細胞株のうち5種 (55.6%) ; SAS,HSC-3,OK92,HO-1-N-1,SCC4でheadpinの発現減弱が認められた。さらに,発現減弱がみられたこれら5種細胞株において4種 (80.0%) ; SAS,HSC-3,OK92,SCC4で脱メチル化剤5-AzaC処理によって発現の回復が確認された。これらの結果はheadpinの発現減弱が口腔扁平上皮癌の進展に関与しており,その機序において過メチル化が関与していることを示唆するものである。
Department of Clinical Molecular Biology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
河崎謙士,鵜澤一弘,倉澤良典,吉田成秀,嶋田 健,上杉尚子,村野彰行,林 幸雄,山木 誠,森谷哲浩,椎葉正史,丹沢秀樹: 口腔扁平上皮癌におけるHeadpin遺伝子 (セリン・プロテアーゼ・インヒビター: serpinファミリー) の発現減弱と過メチル化.
千葉大学大学院医学研究院臨床分子生物学講座
Tel. 043-226-2300. Fax. 043-226-2151. E-mail: m.shiiba@faculty.chiba-u.jp
2008年2月27日受付,2008年4月2日受理.
