




Volume 85, Number 3
doi:10.20776/S03035476-85-3-P113
[Original Paper]
Hideki Hanaoka, Yoshinori Oishi, Akemi Sakamoto,
Kazuhiro Kurasawa, Miki Nishimura, Itsuo Iwamoto
and Yasushi Saito
(Received September 18, 2008, Accepted January 8, 2009)
Natural Killer (NK) T cells regulate immune responses including autoimmunity and are selectively decreased in autoimmune disease. We investigated the regulatory role of CD4- CD8- double-negative (DN) invariant TCR Vα24JαQ T cells, a human counterpart of murine NK T cells, in the development of chronic graft-versus-host disease (GVHD) that resemble to autoimmune disease in clinical features. For this purpose, we monitored frequencies of invariant Vα24JαQ DN T cells in PBL from 8 bone marrow transplantation (BMT) patients, 4 of whom developed chronic GVHD. Invariant Vα24+ DN T cells were recovered at 3 mo. after BMT as a dominated population of DN Vα24 T cells. However, the recovery of these cells was delayed compared with that of conventional T cells. Furthermore, we found that invariant Vα24JαQ DN T cells were not detected at the onset of chronic GVHD and that invariant Vα24JαQ DN T cells were again found after successful treatment for chronic GVHD with prednisolone. These results indicate that the selective reduction of invariant Vα24JαQ DN T cells is related to the disease progression of chronic GVHD, suggesting that the NK T cells may regulate the development of chronic GVHD.
NK T cell, invariant Vα24JαQ, chronic GVHD, regulatory T cell
DN: double negative, TCR: T cell antigen receptor, GVHD: graft versus host disease
Natural killer (NK) T cells have been identified as a novel lymphoid lineage distinct from conventional T cells and NK cells. NK T cells express both T cell receptor (TCR) and NK1 antigen, a member of the family of NKR-P1 natural killer cell receptors[1-4]. NK T cells are CD4- CD8- double negative (DN) or CD4 positive T cells in mice[1-4]and DN T cells in human[5-8]. These NK T cells have unusual features in comparison with conventional T cells and may play an important role in the regulation of some immune responses. First, NK T cells use highly restricted TCR. Murine NK T cells use an invariant TCR Vα14Jα281 that preferentially pairs with Vβ8.2, Vβ7, and Vβ2[9,10]. Human NK T cells are DN T cells bearing invariant Vα24JαQ that has a high homology with murine Vα14Jα281 chain in both the amino acid and nucleotide sequences[5-8]and are paired with Vβ11 and Vβ13 which also have a high homology with murine Vβ8 and Vβ7[7,8]. This highly restricted TCR on NK T cells recognizes a monomorphic MHC class I-like molecule CD1, rather than polymorphic MHC molecules[11-15]. Second, NK T cells can promptly produce large amounts of interleukin-4 (IL-4) and interferon-γ (IFN-γ) by stimulation with anti-CD3 antibody[15-20]. In addition, NK T cells have cytolytic activities in Fas/FasL-[21]and perforin/granzymes- mediated manners[22]. Third, NK T cells have been suggested to regulate the autoimmune process[23,24]. NK T cells are decreased in correlation with the disease activity in autoimmune diseases in mice[23-25]. Furthermore, selective reduction of invariant Vα24JαQ DN T cells (human NK T cells) has been reported in human autoimmune diseases including systemic sclerosis, diabetes and systemic lupus erythematosus (SLE)[26-28]. Thus, NK T cells are importantly involved in regulation of immune reactions including immune diseases.
Chronic graft-versus-host disease (GVHD) is immune disease caused by anti-host immune responses and is the most frequent delayed complication after bone marrow transplantation[29,30]. Clinical features of chronic GVHD were similar to those in autoimmune diseases[30-32]. Chronic GVHD causes scleroderma similar to systemic sclerosis. Chronic GVHD also develops liver injury resembling to autoimmune hepatitis or primary biliary cirrhosis. Keratoconjunctivitis and sialoadenitis like in Sjogren syndrome are developed in chronic GVHD. In addition, esophageal strictures, intestinal involvement and pulmonary insufficiency similar to that in systemic sclerosis were observed in chronic GVHD. Furthermore, autoantibodies were detected in sera from chronic GVHD patients. Thus, clinical features similar to autoimmune diseases and the presence of autoantibodies in chronic GVHD suggest that immunological mechanisms similar to that in autoimmune diseases play important roles in the development of chronic GVHD. However, it remains to be clarified the regulatory role of invariant Vα24JαQ DN T cells (human NK T cells) in the development of chronic GVHD.
Therefore, to elucidate this issue, we analyzed the frequency of invariant Vα24JαQ DN T cells in recipients with BMT every month after BMT, and at the onset of chronic GVHD and after prednisolone therapy. Our results showed that invariant Vα24JαQ DN T cells were recovered by 3 mo. after BMT. Moreover, our results indicate that invariant Vα24JαQ DN T cells are closely related to the disease progression of chronic GVHD, as indicated by the selective reduction of DN Vα24JαQ T cells at the onset of and during chronic GVHD and the recovery of those cells after successful prednisolone therapy, suggesting that NK T cells might be involved in the development of chronic GVHD.
Fourteen adult recipients enrolled in this study gave informed consent in accordance with the Declaration of Helsinki, and an ethics committee at site approved the protocol. Recipients were diagnosed as leukemia, lymphoma or aplastic anemia and were given allogenic BMT at Chiba university hospital in 1997. All adult recipients were studied at every 1 mo. after BMT and at the onset of chronic GVHD. Six recipients dropped out from this study because of BMT failure and of relapse. We evaluated data from eight recipients (3 males and 5 females, aged 19 to 34) included in this study who had not dropped out from this study. Our data from Chiba university BMT recipients (not published) shows chronic GVHD develops 40-60% after BMT and in this study four recipients were free from chronic GVHD and four recipients were not. Eight adult recipients who were diagnosed as leukemia, lymphoma or aplastic anemia and were given allogenic BMT (3 males and 5 females, aged 19 to 34) were studied at every 1 mo. after BMT and at the onset of chronic GVHD. All recipient were prepared with cyclophosphamide (60 mg/kg for 2 days) followed by total-body irradiation (12 Gy) before BMT. After total-body irradiation, 3x108 cells/kg of bone marrow cells from unrelated donors were infused intravenously. G-CSF was used during the period of granulocytopenia for 7 to 14 days. To prevent development of acute GVHD, 100 to 150 mg/day cyclosporine was used for 28 days and methotrexate was given for four times (15 mg/㎡ on day 1, 10 mg/㎡ on day 3, 6, 11 after BMT).
Acute GVHD was diagnosed based upon clinical findings and histopahological confirmation when possible and graded as 1994 Consensus Conference[33]. Six of eight patients (case 3-8) developed acute GVHD by 1mo after BMT. One patient (case 3) had developed grade III GVHD involving skin and intestine. Other five patients developed grade I GVHD involving skin in five patients and liver in one patient. All acute GVHD patients were treated with prednisolone (2 mg/kg/day) for 14 days following reduction of dosage and their symptoms were improved. No symptoms and abnormal findings were observed at 3 mo after BMT.
Chronic GVHD was diagnosed based upon clinical findings and histopahological confirmation when possible and was graded as criteria by Shulmann et al[34]. Four out of eight recipients (case 1-4) developed chronic GVHD. One patient (case 2) developed limited GVHD with skin involvement and three patients developed extensive type GVHD involving skin and liver (case 1 and 4) and skin and lung (case 3). All patients with chronic GVHD were treated with prednisolone at the initial dose of 2 mg/kg/day for 2 to 6 weeks, and then the dosage was gradually reduced. At 6 mo. after BMT, all patients were improved and no symptoms and abnormal findings were observed after 2 mo. prednisolone therapy. Four healthy subjects, sex and age matched, were also examined as controls.
Peripheral blood lymphocytes (PBL) were isolated from 50 ml of heparinized peripheral venous blood of eight BMT recipients and five healthy subjects by Ficoll-Paque (Pharmacia Fine Chemicals, Uppsala, Sweden) density gradient centrifugation. Cells (1 x 106) were stained with fluorescence- or biotin-conjugated antibodies in phosphate-buffered saline containing 1 % fetal calf serum for 30 min at 4 ℃. The following fluorescein isothiocyanate (FITC)-, phycoerythrin (PE)-, or biotin-conjugated monoclonal antibodies (mAb) were used: CD4 (Leu-3a), CD8 (Leu-2a), TCR αβ (Becton Dickinson, Mountain View, CA) and TCR Vα24 (Cosmo Bio Co., Tokyo, Japan). Cells stained with biotinylated mAb were then incubated with streptoavidin-PE or -Tricolor (Caltag, San Francisco, CA). Stained cells were resuspended in phosphate-buffered saline containing 1 % fetal calf serum and analyzed by FACScan (Becton Dickinson) using Cell Quest program.
CD4- CD8- double-negative (DN) TCR αβ T cells were sorted from PBL of BMT recipients and healthy subjects by FACStar (Becton Dickinson) using anti-CD4 plus anti-CD8 mAb. The yields of DN T cells were approximately 1 x 105.
Total RNA (0.1-1 μg) was prepared from sorted DN T cells by the method of acid guanidinium thiocyanate/phenol/chloroform extraction using Isogen solution (Nippon Gene Co., Tokyo, Japan). The first strand complementary DNA (cDNA) was then synthesized from 0.1-1 μg of total RNA in 20 μl of reaction buffer containing oligo-dT primer using avian myeloblastosis virus reverse transcriptase. The reaction mixture was incubated at 25 ℃ for 10 min and then at 42 ℃ for 60 min.
To amplify invariant Vα24JαQ gene, we used nested polymerase chain reaction (PCR) method. cDNA from DN T cells were first amplified by PCR using primers for Vα24 (5-AAGCAAGATACTGGGAGAGGT-3) and Cα (5-ATGTCTAGCACAGTTTTGTCTGTG-3). The denaturing step was done at 95 ℃ for 1.5 min, the annealing step at 60 ℃ for 1 min, and the extension step at 72 ℃ for 1 min, for 25 cycles on a DNA thermal cycler (Perkin-Elmer Corp., Norwalk, CT). PCR products were then amplified by polymerase chain reaction (PCR) using primers for invariant Vα24JαQ (5-AGCGACAGAGGCTCAACCCT-3) and Cα (5-GGTGAATAGGCAGACAGACTT-3) at 95 ℃ for 1.5 min, the annealing step at 62 ℃ for 1 min, and the extension step at 72 ℃ for 1 min, for 25 cycles. For detection of Vα24 gene products, first PCR products were then amplified using inner Vα24 primer (5-CTCAGCGATTCAGCCTCCTAC-3) and inner Cα primer (5-GGTGAATAGGCAGACAGACTT-3) for 25 cycles. The PCR products were loaded at 2% agarose gel and detected by ethidium bromide (Nippon Gene, Co. Ltd.). This nested PCR method allowed us to detect one invariant Vα24JαQ T cells in 1-5 x 105 cells.
TCR Vα24 cDNAs from DN T cells were amplified by PCR using primers for Vα24 with an EcoRI restriction site (5’-CGAATTCCTCAGCGATTCAGCCTCCTAC-3’) and Cα (5’-CGAATTCGGTGAATAGGCAGACAGACTT-3’). The denaturing step was done at 95 ℃ for 1.5 min, the annealing step at 60 ℃ for 1 min, and the extension step at 72 ℃ for 1 min, for 30 cycles on a DNA thermal cycler. PCR products were purified by phenol extraction, precipitated with ethanol, and digested with excess amounts of EcoRI. The DNA fragments with expected sizes of the cDNAs were enriched by preparative low-melting-point agarose gel electrophoresis. The recovered DNA fragments were ligated to M13mp19 plasmids obtained by EcoRI digestion. Phages were grown on TG-1 Escherichia coli cells and recombinant phage DNA was purified for colony PCR. DNA from each colony was amplified using primers for invariant Vα24JαQ and Cα and those for Vα24 and Cα for detection for invariant Vα24JαQ and Vα24 genes, respectively as mentioned above. To confirm the results by colony PCR, some clones including positive and negative for Vα24JαQ gene by colony PCR were sequenced by the dye primer method using an automated sequencer (Applied Biosystems). The results of sequence analysis were consistent with those by colony PCR.
Data are summarized as mean ± SD. The statistical analysis of the results was performed by the unpaired and paired t-test. P values <0.05 were considered significant.
To determine whether NK T cells were recovered after BMT, we analyzed invariant Vα24JαQ DN T cells in PBL from 8 BMT recipients every month after BMT. For this purpose, we used nested PCR method to detect the invariant Vα24JαQ gene in which Vα24 genes are amplified in 1st PCR and then invariant JαQ gene was expanded in second PCR. This method could detect one Vα24JαQ bearing cell in 1-5 x 105 cells (data nit shown).
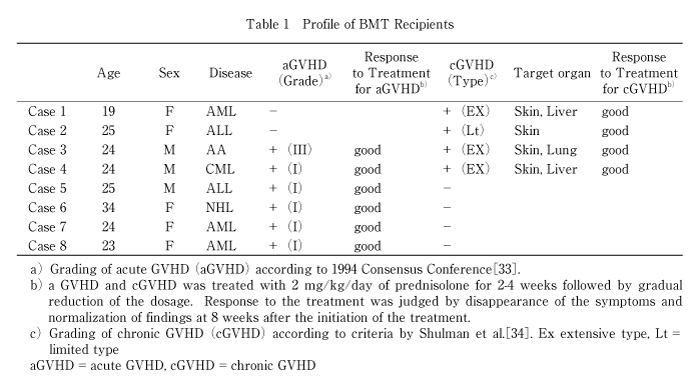
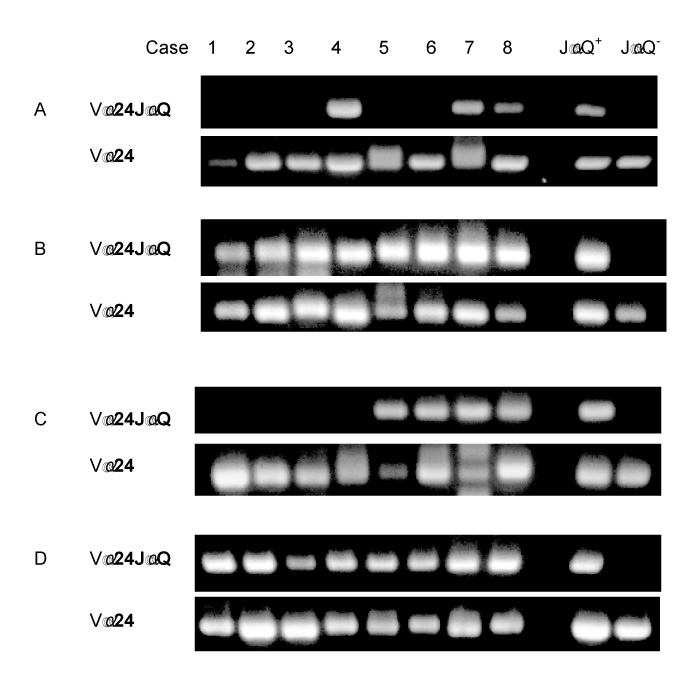
Fig. 1
Expression of invariant Vα24JαQ genes in DN T cells in PBL from BMT recipients
Invariant Vα24JαQ mRNA expression was examined by nested PCR in DN T cells from PBL of BMT recipients at 1 mo. after BMT (A), and at 3 mo. after BMT (B). Invariant Vα24JαQ mRNA expression was examined by nested PCR in DN T cells from PBL of BMT recipients at the onset of chronic GVHD (4 mo. after BMT) (C), and after prednisolone therapy for GVHD (6 mo. after BMT) (D). Case 1-4 developed chronic GVHD at 4 mo. after BMT. These patients were successfully treated with prednisolone (2 mg/kg for 2-6weeks followed by decrease of dosage) and no symptoms and abnormal findings were observed at 6 mo. after BMT. Fig. 2 Cell number of DN invariant Vα24JαQ T cells in BMT recipients with chronic GVHD DN invariant Vα24JαQ T cells was assessed in four BMT recipients without chronic GVHD (left) and with chronic GVHD (right) before the onset of chronic GVHD (at 3 mo. after BMT), at the onset of chronic GVHD (at 4 mo. after BMT), and after successful prednisolone therapy for GVHD (at 6 mo. after BMT).
In healthy controls, invariant Vα24JαQ T cell is a dominant population in DN Vα24 T cells[5-8]and the frequencies of invariant JαQVα24 T cells in DN Vα24 T cells were 50-100% (mean ± sd; 84.1 ± 16.3%) in our study. To determine whether recovered invariant Vα24JαQ T cells are major population in DN Vα24 T cells, we examined the frequencies of invariant Vα24JαQ gene in Va24 clones in peripheral blood DN T cells of BMT recipients. Invariant JαQ gene was dominant in Vα24 clones in DN T cells of all BMT recipients at a high frequency (5/16 to 16/16, mean ± SD: 84 ± 16%) (Table 2).
Thus, recovered invariant Vα24JαQ DN T cells are a major population in DN Vα24 T cells as observed in healthy controls.
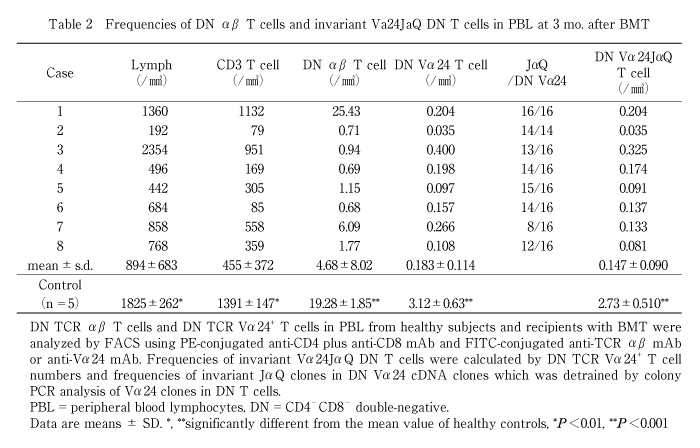
We then estimated absolute numbers of invariant Vα24JαQ DN T cells based on lymphocytes number, percentage of DN Vα24 T cells and frequencies of invariant Vα24JαQ clones among Vα24 clones in DN T cells. The percentage of recovered cell numbers to control cell numbers was smaller in invariant Vα24JαQ DN T cells than in conventional T cells. At 3 month after BMT, lymphocyte number, T cell number, and DN T cells in BMT recipients were smaller than those in healthy controls and were about 50%, 30%, and 25% of healthy controls, respectively (Table 2). Cell numbers of DN Vα24 T cell and invariant Vα24JαQ DN T cell was about 6% and 5% of healthy controls, respectively (Table 2).Thus, recovery of NK T cells after BMT was impaired and delayed compared to that of conventional T cells.
These results indicate that NK T cells were recovered by 3 mo. after BMT and were a major population of DN Vα24 T cells, although recovery of NK T cells was delayed compared to conventional T cells.
Clinical features of chronic GVHD resemble those of autoimmune diseases such as systemic sclerosis[30,32,35,36]. In autoimmune diseases such as systemic sclerosis, SLE, NK T cells, invariant Vα24JαQ NK T cells, were decreased[26-28]. Therefore, to determine whether DN invariant Vα24JαQ T cells were decreased in chronic GVHD, we analyzed the invariant Vα24JαQ gene in DN lymphocytes by PCR method in recipients with chronic GVHD before prednisolone therapy. Four patients (case 1-4) developed chronic GVHD at 4 months after BMT.
At the onset of chronic GVHD, invariant Vα24JαQ DN T cells was decreased. Invariant Vα24JαQ gene was detected in DN lymphocytes from all recipients at 3 mo. after BMT before the onset of chronic GVHD (Fig. 1B). At 4 mo. after BMT when chronic GVHD was developed, invariant Vα24JαQ gene was not detected in all four patients who developed chronic GVHD (Fig. 1C). In contrast, the invariant Vα24JαQ gene was detected in DN lymphocytes from BMT recipients who did not develop chronic GVHD. In addition, disappearance of invariant Vα24JαQ DN T cells was observed during the active chronic GVHD. Thus, invariant Vα24JαQ DN T cells were disappeared in chronic GVHD.
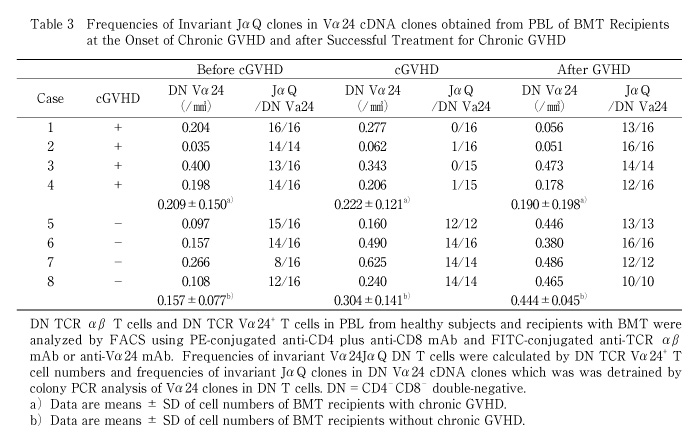
We also examined frequencies of invariant Vα24JαQ gene in Vα24 clones in cGVHD patients and estimated the cell numbers. The absolute number of invariant Vα24JαQ DN T cells was decreased in all four BMT recipients when they developed chronic GVHD (before c-GVHD 0.184 ± 0.119/mm3 vs. in c-GVHD 0.011 ± 0.020/mm3, P<0.05), although DN Vα24 T cell number did not decreased (before c-GVHD 0.209 ± 0.150/mm3 vs. in c-GVHD 0.222 ± 0.121/mm3) (Table 3 and Fig. 2).
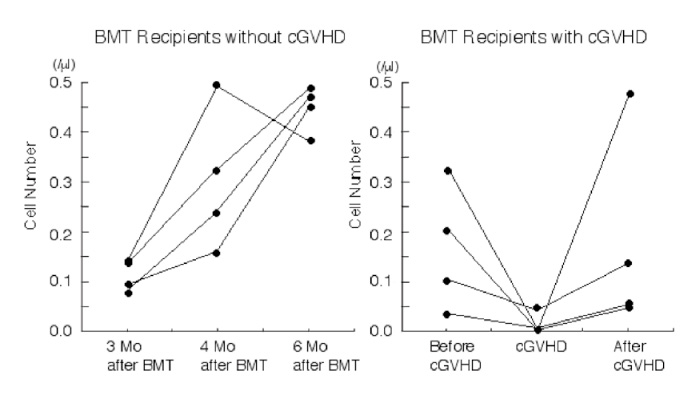
Fig. 2
Cell number of DN invariant Vα24JαQ T cells in BMT recipients with chronic GVHD
DN invariant Vα24JαQ T cells was assessed in four BMT recipients without chronic GVHD (left) and with chronic GVHD (right) before the onset of chronic GVHD (at 3 mo. after BMT), at the onset of chronic GVHD (at 4 mo. after BMT), and after successful prednisolone therapy for GVHD (at 6 mo. after BMT).
In contrast to chronic GVHD patients, invariant JαQ gene was dominantly detected at high frequencies (14/16 to 16/16 or 14/14) in BMT recipients who did not develop chronic GVHD at 4 mo. after BMT. The absolute number of invariant Vα24JαQ was increased in BMT recipients without chronic GVHD (3 mo. 0.111 ± 0.029/mm3 vs. 4 mo. 0.304 ± 0.141/mm3, P<0.01) (Table 3, Fig. 2).
Thus, invariant Vα24JαQ DN T cells are selectively reduced in chronic GVHD and no clonal expansion of DN Vα24 T cells bearing TCR other than invariant Vα24JαQ gene was observed.
Recovery of invariant Vα24JαQ DN T cells by prednisolone therapy in chronic GVHD patients
To determine whether Vα24JαQ T cells are related to the disease activity of chronic GVHD, we then examined invariant Vα24JαQ gene by PCR method in DN lymphocytes from the same four chronic GVHD patients who were treated with prednisolone and chronic GVHD was rendered inactive states.
In inactive chronic GVHD, invariant Vα24JαQ gene was again detected in DN lymphocytes from all four chronic GVHD patients treated with prednisolone (Fig. 1D). Furthermore, colony PCR analysis of Vα24 clones of DN T cells revealed that invariant Vα24JαQ T cells dominated in all four patients (Table 3). In two patients (case 2 and 3), all clones were invariant Vα24JαQ. In other two patients (case 1 and 4), invariant JαQ gene also dominated in DN Vα24+ T cells at a frequency of 13/16, and 12/16, respectively. The absolute number of invariant Vα24JαQ T cells was increased after prednisolone therapy in all four chronic GVHD patients (before prednisolone therapy 0.011 ± 0.020/mm3 vs. after prednisolone therapy 0.176 ± 0.202, P=0.10) (Fig. 2). Thus, invariant DN T cells were reappeared after prednisolone therapy and the reduction of these invariant Vα24JαQ DN T cells were correlated with disease activity of chronic GVHD.
In this study, we showed that invariant Vα24JαQ DN T cells, human NK T cells, were recovered by 3 mo. after BMT and that the recovery of invariant Vα24JαQ DN T cells was delayed compared to that of conventional T cells. Moreover, we demonstrated that invariant Vα24JαQ DN T cells are related to the disease progression of chronic GVHD, as indicated by selective reduction of invariant Vα24JαQ T cells in chronic GVHD and the recovery of those cells after successful prednisolone treatment for chronic GVHD, suggesting that NK T cells might be involved in the development of chronic GVHD.
Many studies about immune reconstitution after BMT have shown that increase of NK cells was first found during first 2 mo. after BMT and then T cells were gradually increase after sharp increase of NK cells and normalization of T cell numbers takes 6- 24 mo[37-39]. However, the recovery of NK T cells after BMT had been unknown. Here, we first demonstrated that NK T cells appear by 3 mo. after BMT.
It is still unknown whether invariant Vα24JαQ DN T cells found at 3 mo. after BMT were originated from donor or host. In this study, we failed to sort invariant Vα24JαQ DN T cells to decide the origin, because these cells are too small population for sorting at 3 mo. after BMT. However, we believe that these invariant Vα24JαQ T cells were donor origin, because 1) Fluorescence in situ hybridization (FISH) analysis for sex chromatin and PCR analysis for variable number of tandem repeats (VNTR) revealed that leukocytes were 100% donor origin at 3 mo. after BMT, 2) invariant Vα24JαQ DN T cells continued to increase from 3 mo. after BMT (Fig. 2). Furthermore, invariant Vα24JαQ DN T cells at 3 mo. after BMT expressed CD45RA, a naive T cell marker, and did not bear CD45RO, a memory T cell marker, that are expressed on NK T cells in healthy controls even in cord blood cells[40](our unpublished data), suggesting that those cells were immature NK T cells probably originated from donor stem cell origin rather than from remaining host cells or contaminated donor NK T cells.
We demonstrated that recoveries of invariant Vα24JαQ DN T cells are delayed compared with that of conventional T cells. CD1d recognition is essential for NK T cell development, which is shown by defective NK T cells in CD1 deficient mice[12-14]. Since CD1d is expressed on dendritic cells[4,15,22], NK T cells development after BMT might occurre after the reconstitution of CD1d positive dendritic cells. This might explain the delay of reconstitution of NK T cells. In addition, NK T cells required also IL-7 and IL-15 for their development[41,42]. Impaired cytokine microenvironment after BMT might also delay the development of NK T cells. Moreover, if NK T cells are originated from donor stem cell and require thymic environment for their development, delayed or impaired reconstitution is not surprising, because it has been shown that majority of recovered conventional T cells are memory T cells and that generation of naive T cells from stem cells were impaired and takes long time in adult BMT recipients[43,44].
In this study, we demonstrated selective reduction of invariant Vα24JαQ DN T cells in chronic GVHD. We previously demonstrated a selective reduction of invariant Vα24JαQ DN T cells in patients with systemic sclerosis[26]. In addition, decrease of NK T cells has been reported in autoimmune diseases such as type I diabetes[27]. Moreover, we recently found that invariant Vα24JαQ DN T cells were decreased in active SLE and reappeared after prednisolone therapy[28]. Therefore, decrease of invariant Vα24JαQ DN T cells in chronic GVHD suggests that similar immunological abnormalities exist in autoimmune diseases and chronic GVHD as well as clinical features.
Dysregulation of cytokine production has been shown in chronic GVHD. In murine chronic GVHD, T cells with Th2 type cytokine profiles are activated and IL-10 production is enhanced[45]. In human chronic GVHD, cytokine dysregulation was also observed[46,47]and serum IL-10 levels were elevated[48]. Furthermore, IL-10 has been shown to down regulate CD1 molecules, critical molecules for NK T cells development and activation, on monocytes[49]. Therefore, increased production of IL-10 in chronic GVHD may account for the decrease of DN Vα24JαQ T cells. Indeed, we found that the cell growth of DN Vα24JαQ T cells from healthy subjects was significantly inhibited by the addition of IL-10 into the culture (our unpublished data). Interestingly, increased IL-10 production was found in SLE[50,51]and systemic sclerosis patients[52]in whom invariant Vα24JαQ DN T cells were selectively decreased, suggesting that cytokine dysregulation including IL-10 may be shared mechanism for reduction of NK T cells.
The pathophysiologic role of NK T cells in chronic GVHD is to be elucidated. The roles of NK T cells in modulating autoimmune diseases have been suggested, of which clinical features and immunological abnormalities were similar to chronic GVHD. Mieza et al[24]demonstrated that in vivo deletion of NK T cells accelerated the development of autoimmune disease in lpr mice, lupus-prone mice, and the introduction of Vα14Jα281 transgene delayed the onset of the disease, suggesting that NK T cells play a role in regulating auto reactive T cells in the autoimmune process. Recently, Ikehara et al[53]reported that NK T cells plays an critical roles in acceptance of xenograft by cytokine independent manner. Thus, NK T cells could serve as immunosuppressive regulatory cells and could regulate the development of immunological diseases including autoimmune diseases and chronic GVHD. The loss of NK T cells might accelerate the development of chronic GVHD.
The regulatory roles by NK T cells in chronic GVHD might be mediated by cytokines produced by NK T cells. NK T cells produce large amounts of IL-4 and IFN-γ upon stimulation. IL-4 produced by NK T cells suppresses the development and effects of Th1 type cells[54]. It has been shown that IL-4 producing NK T cells suppressed acute GVHD that is mediated by Th1 cells in murine model[55]. On the contrary, IFN-γ regulates Th2 cells[56]that contribute to the development of chronic GVHD[45-47]. Indeed, we found large amounts of IFN-γ production by invariant Vα24JαQ DN T cells in healthy controls and BMT recipients at 3 mo. after BMT (our unpublished data). Thus, cytokines produced by NK T cells might regulate the development of chronic GVHD.
In summary, we have shown that invariant Vα24JαQ DN T cells was reconstituted after BMT, which was delayed compared with conventional T cells and that the selective reduction and recovery of invariant Vα24JαQ DN T cells is related to the disease progression in chronic GVHD.
This work was supported in part by grants from the Ministry of Education, Science and Culture and from the Ministry of Health and Welfare, Japan.
【目的】マウスNK T細胞はIL-4,IFN-γを産生し,免疫系の調節細胞と考えられている。この細胞はアロ反応性を抑制し,GVHDを抑制する細胞である可能性が指摘されている一方で,GVHDにて細胞傷害活性を発揮し病態増悪因子であることも報告され,未だGVHDにおけるNK T細胞の果たす役割は明確にされていない。本研究ではヒトNK T細胞と考えられるCD4-8-(DN) Va24 JaQ T細胞(NK T細胞)が慢性GVHD(cGVHD)の病態と関連する可能性があるか否かを検討する目的で,骨髄移植(BMT)後の末梢血中のNK T細胞が再構築する時期,NK T細胞とcGVHD発症との相関について解析を行った。
【方法】 BMT症例8例(BMT後cGVHD発症4例,非発症例4例)の末梢血を用いた。cGVHD発症前後でのNK T細胞のflow cytometry法,RT-PCR法等による量的解析を行った。
【結果】BMT症例8例のNK T細胞は末梢血単核球中の0.02-0.04%であり健常人と比較し低値であり経時的に変動は認められなかった。このうちNK T細胞の再構築をRT-PCR法によりBMT後1から3ヶ月に認めた。cGVHD発症群では発症時にNK T細胞が減少し,ステロイド治療による症状改善と共にNK T細胞は再増加した。一方,cGVHD非発症群では全経過においてNK T細胞が検出された。
【結論】NK T細胞はBMT後3ヶ月で再構築された。このNK T細胞はcGVHDの活動性に相関して減少,消失することからNK T細胞はcGVHDの病態に関与している可能性が示唆された。
Clinical Cell Biology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
花岡英紀,大石嘉則,坂本明美,倉沢和宏,西村美樹,岩本逸夫,齋藤 康: 慢性GVHDにおけるNK T細胞の役割.
千葉大学大学院医学研究院細胞治療学
Tel. 043-226-2630. Fax. 043-226-2629. E-mail: hanaoka.hideki@mac.com
2008年12月25日受付,2009年3月18日受理.
