




Volume 87, Number 2
doi:10.20776/S03035476-87-2-P61
[Original Paper]
Muhammed Zahed1),*, Toshikazu Suzuki1,2),*, Kazuko Kita1)
Shigeru Sugaya1),* and Nobuo Suzuki1),*
(Received November 18, 2010, Accepted December 20, 2010)
Increased expression of glucose-regulated protein 94(GRP94)on the cell surface is associated with tumorigenicity, suggesting that small peptides bound to GRP94 may be useful for drug delivery targeted to cancer expressing cell surface GRP94 . In this study, candidate peptides that exhibited high affinity binding to the carboxy-terminal fragment of the GRP94 (GRP94-C) protein were screened using a phage display library. Three high affinity peptides (No. 6; CTLMSDPAC, No. 58; CSLVPIPAC, No. 72; CQTKPLFQC)were identified. Kd values for all three peptides binding to target GRP94-C protein were within the range of 0.39-0.46 uM. These novel peptide sequences could potentially be used as a part of a peptide-conjugated drug system, designed to selectively target cancer cells with surface expression of GRP94.
phage display, GRP94, QCM analysis
Peptide ligands have attracted attention as powerful molecular tools in the treatment of cancer patients[1,2]. Current chemotherapy using standard anticancer drugs often leads to severe side effects, causing extreme distress to the patient. Development of methodologies that allow for cancer cell-specific delivery of therapeutic agents offers a potential strategy in decreasing these harmful and undesirable side effects[3,4].
Recent studies have shown that peptide ligands bound to glucose-regulated protein 78 (GRP78)can successfully be used for peptide-based drug delivery[5]. GRP78, a member of the HSP70 family, is an endoplasmic reticulum (ER)protein that possesses a carboxy(C)-terminal KDEL sequence that constitutes a feedback signal from the Golgi to the ER[6]. GRP78 is principally located in the ER; however, reports also indicate its existence on the cell surface. Expression levels in the ER and on the cell surface are both increased under stress conditions[7,8], and high expression levels on the cell surface are linked to a greater degree of cancer malignancy [9,10].
GRP94, a paralog of HSP90, is also an endoplasmic reticulum(ER)protein that possesses peptide and adenine nucleotide-binding domains. Like other proteins residing in the ER, GRP94 contains a C-terminal KDEL sequence. GRP94 also exists on the cell surface and functions as an antigen-representing chaperon protein[11]. Increased expression of GRP94 on the cell surface has also been demonstrated to be associated with tumorigenicity[12,13].
In this study, we focused on the discovery of GRP94-binding peptides. Since the amino (N)-terminal fragment of the GRP94 protein is involved in peptide presentation via professional antigen presenting cells[14], it is possible that any peptide can bind to this fragment sequence-independently. We therefore searched for peptides that could bind to the C-terminal fragment of the GRP94 protein.
Full-length GRP94 cDNA(accession no. X15187) was amplified using a forward primer(5'-AAA GGA TCC ATG AGG GCC CTG TGG GTG CT-3') and a reverse primer (5'-AAA GAA TTC TTA TTC AGC TGT AGA TTC CT-3') and inserted into the pGEX-6P-1-GST fusion protein expression vector (GE Healthcare UK Limited, Buckinghamshire, UK) to yield pGEX-GRP94. The plasmid was further digested with BamH1 and HindIII, and a 3.7-kb fragment containing the C-terminal domain of GRP94(amino acids 493 to 799) was re-inserted into the empty expression vector pGEX-6P-1 to yield pGEX-GRP94-C.
BL21 competent cells were transformed with pGEX-GRP94-C. Transformants were cultured in 2×YT medium containing 100 ug/ml ampicillin at 30℃, and fusion protein expression was induced using isopropyl-1-thio-Β-D-galactopyranoside (IPTG). Glutathione S-transferase (GST) and the C-terminal domain of the GRP94 fusion protein (GST-GRP94-C) was bound to a GSTrap FF column (GE Healthcare UK Limited) and further digested overnight with PreScission Protease (GE Healthcare Limited, UK) at 4℃. The C-terminal domain of GRP94 (~37.2 kDa-peptide; GRP94-C) was subsequently eluted with a buffer without glutathione.
Peptide sequences were selected from a commercially available disulfide constrained 7-mer random library displayed on phage M13 via N-terminal fusion to the minor coat protein, g3p, with a diversity of 1.3×109 (Ph. D.-C7C™ phage display peptide library kit, New England Biolabs, Beverly, MA, USA), as described previously[15]. Initially, recombinant GRP94-C protein was biotinylated using a sulfo-NHS-LC-biotin (Pierce, USA) according to the manufacturer’s instructions. Biotinylated protein (0.1 ug) and 2×1011 phage particles from the Ph. D.-C7CTM phage display peptide library kit were mixed in 400 ul of Tris-buffered saline (50mM Tris pH 7.5, 150mM NaCl) supplemented with 1% (v/v) Tween-20 (TBS-T) to prepare the phage-target complex. Phage-target complex was then panned against a streptavidin-coated plastic plate for affinity capture. After washing, bound phages were eluted with 1mM dithiothreitol (DTT) and amplified for next-round panning. After a third round of amplification and panning for binding GRP94-C protein, individual phages were isolated and the displayed peptide sequences were deduced after DNA sequencing.
Candidates were selected for the ELISA assay based on amino acid similarity. Ninety-six-well microtiter plates (Nunc Maxisorp, Denmark) were coated with 200 ul per well of GRP94-C protein (100 ug/ml). 0.1 M NaHCO3 (pH 8.6) and incubated overnight at 4℃. After washing with TBS-T, wells were incubated with filter-sterilized blocking buffer (0.1 M NaHCO3 pH 8.6, 5mg/ml bovine serum albumin, 0.02% NaN3) to reduce any non-specific interaction of phages to the plate. Fourfold-serially diluted samples of selected phages were then placed in ELISA microplates and incubated for 2 h at room temperature. After the unbound phages were removed by washing with TBS-T, a horseradish peroxidase (HRP)-conjugated anti-M13 antibody (GE Healthcare Limited) diluted with blocking buffer at a ratio of 1:5000 was added to the plates and incubated for 1h at room temperature. Immune complexes were then detected using 2, 2'-azino-di-[3-ethylbenzthiazoline sulfonate] diammonium salt (Roche, Berlin, Germany) as a substrate for HRP. After a 20-min incubation at room temperature, the absorbance at 405 nm was determined using an automated ELISA reader (Model E-max, Molecular Devices, CA, USA). From the pool of candidate peptides (Table 1), a single randomly selected peptide (peptide No.34) with a lower or no amino acid similarity to the above selected peptides was used as a negative control under identical experimental conditions.
Cysteine-constrained peptides with biotinylated amino termini were purchased from Scrum Co. Ltd. (Tokyo, Japan). Intra-molecular disulfide bridge formation of the peptides was characterized using matrix-assisted laser desorption/ionization time-of-flight mass spectroscopy before and after reduction using 1mM DTT and also by their unreactivity to Ellman’s reagent, a sulfhydryl assay reagent (Pierce, IL, USA).
Binding analysis of the interaction between GRP94-C and synthetic peptides was performed with a 27 MHz QCM machine (Affinix Q4, Tokyo, Japan). Briefly, the sensor chip was initially cleaned twice with 1% SDS solution and rinsed with phosphate-buffered saline (PBS, 150 mM NaCl in 10 mM phosphate buffer, pH 7.2). Neutravidin (10 ug/ml) was then absorbed onto the chip for immobilization of biotinylated peptides. Biotinylated peptide solution (10 uM) was further added to the neutravidin-absorbed sensor chip for one hour at room temperature. Finally, GRP94-C protein was added to the peptide-immobilized chip at different concentrations for binding measurements and the Kd value was calculated. Purified GST protein was used as a negative control under identical experimental conditions.
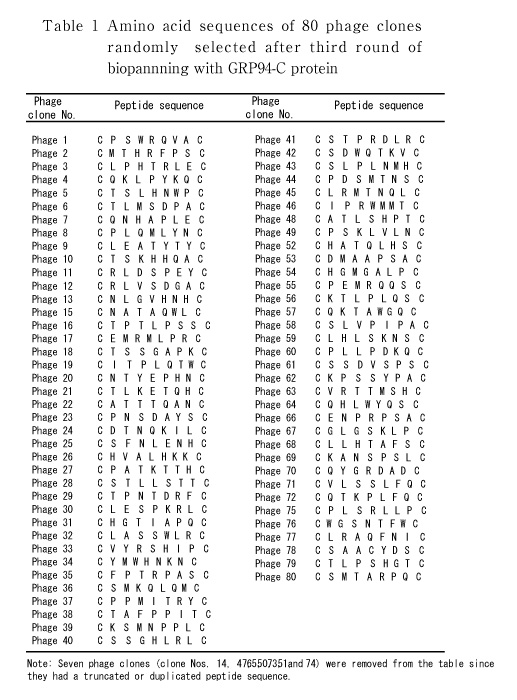
To screen peptide ligands for the GRP94-C protein, we used a phage display peptide library displaying 7-mer cysteine constrained random peptides (C-X7-C). During the course of selection, the relative yield of phage recovery improved from 0.000005 (first round) to 0.0001 (third round), which is consistent with enrichment in binding clones. After three rounds of panning using purified GRP94-C protein, 80 phage clones were randomly selected, amplified and sequenced to identify the displayed peptides. Primarily from the pool of 80 candidate peptides (Table 1), 16 different phage peptide sequences were selected and arranged into four groups based on amino acid sequence or consensus motif similarity (Fig. 1).
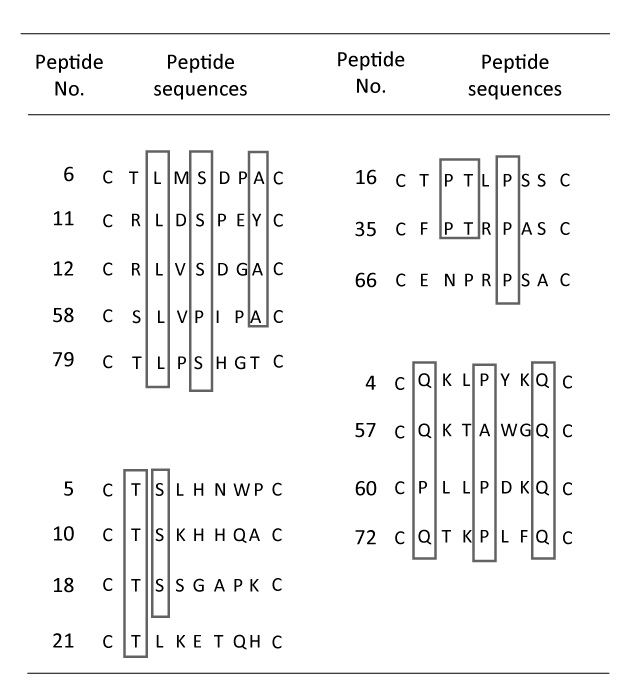
Fig. 1
Grouping of candidates for GRP94-C binding peptides based on amino acid sequence consensus. Peptide sequences were grouped into four groups based on the analogous positions of similar amino acid residues (boxed residues). The four groups are as follows: group 1 (peptide Nos. 6, 11, 12, 58, 79) group 2 (peptide Nos. 5, 10, 18, 21), group 3 (peptide Nos. 19, 35, 66) and group 4 (peptide Nos. 4, 57, 60, 72).
Phage ELISA experiments were performed to test the specific binding of peptides to the target GRP94-C protein. The 16 phages selected(Fig. 1) were individually amplified, serially diluted and incubated with purified recombinant GRP94-C target protein, and a randomly selected phage was investigated in parallel as a negative control. Fourteen phages (excluding phage No. 21 and 35) showed increased absorbance values in the presence of the target protein when compared with the negative control (Fig. 2). We chose eight clones (phage Nos. 4, 6, 18, 58, 60, 66, 72 and 79) that showed the greatest increase in reactivity using the first ELISA. We further performed phage ELISA analyses to confirm the binding strength of phage-borne peptides to GRP94-C protein (Fig. 3). Four of the eight phage clones (phage Nos. 6, 58, 60 and 72) were selected based on their binding activity to GRP94-C and ranked on the basis of the final ELISA results. The best three candidates (phage Nos. 6, 58 and 72, Fig. 4) were chosen for downstream experiments. All three phages showed a similar absorbance range and thus, were expected to bind to GRP94-C with a similar affinity range.
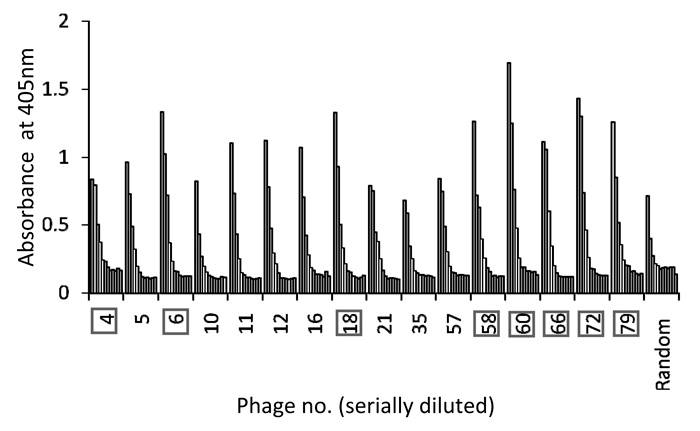
Fig. 2
Result of first ELISA for 16 selected phage clones. Each phage clone was diluted 12 times (fourfold serial dilutions starting with 1012 virions), and absorbance was read at 405 nm by microplate reader. Eight phage clones were selected for the second ELISA experiment (boxed numbers) on the basis of absorbance intensity.
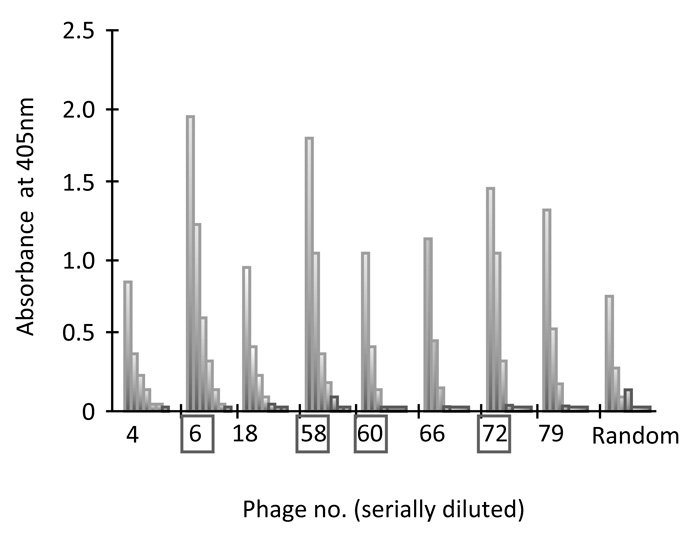
Fig. 3
Result of second ELISA assay for eight selected phage clones. Each phage clone was serially diluted seven times (fourfold serial dilutions starting with 1012 virions), and absorbance at 405 nm was determined using a microplate reader. Four phage clones (boxed numbers) were selected for further ELISA experiments to check reproducibility.
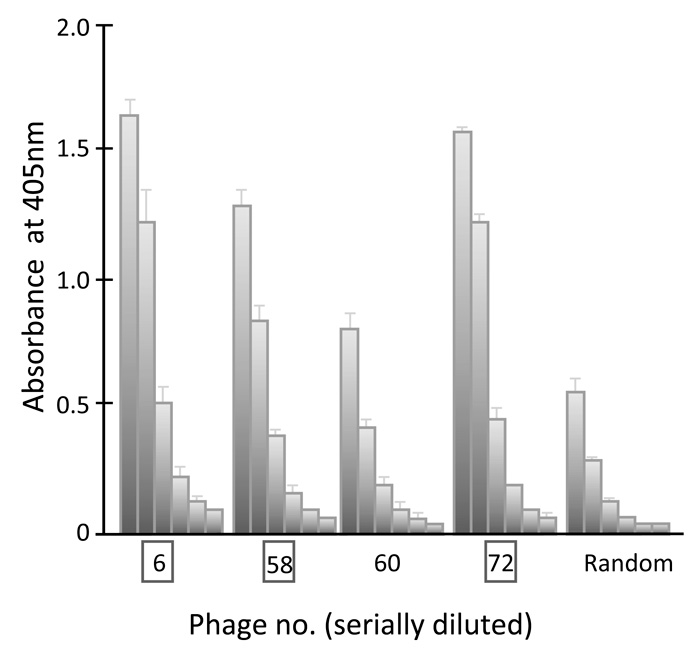
Fig 4.
Interaction of four selected phage clones with GRP94-C analyzed by ELISA. Each phage clone was serially diluted six times (fourfold serial dilutions starting with 1012 virions) and absorbance at 405 nm was determined using a microplate reader. Three phage clones were selected (phage clone Nos. 6, 58 and 72) on the basis of their levels of absorbance.
Binding affinity of the selected three peptides (peptide Nos. 6, 58 and 72) with GRP94-C protein was measured by QCM analysis. Protein binding to the immobilized peptide was calculated based on the Sauerbrey equation[16], which represents a linear dependence between frequency shift and the bound GRP94-C protein. Binding of the target GRP94-C protein to the peptides immobilized on the QCM sensor chip was observed as a decreased frequency (Fig. 5A). Addition of purified GST protein did not produce a shift in frequency and suggested that the GST proteins did not bind to the immobilized peptides (Fig. 5A). The affinity constant (Kd) value was determined by plotting absorbed GRP94-C protein in terms of frequency shift (ΔF) versus the concentration of injected GRP94-C in the buffer solution (Fig. 5B). Kd values for all three peptides (peptide Nos. 6, 58 and 72) binding to target GRP94-C protein were within the range of 0.39-0.46 uM (0.45 uM, 0.46 uM and 0.39 uM, respectively). No significant difference in the binding affinity of these peptides to GRP94-C was observed, although peptide No. 72 did show a slightly higher binding affinity.
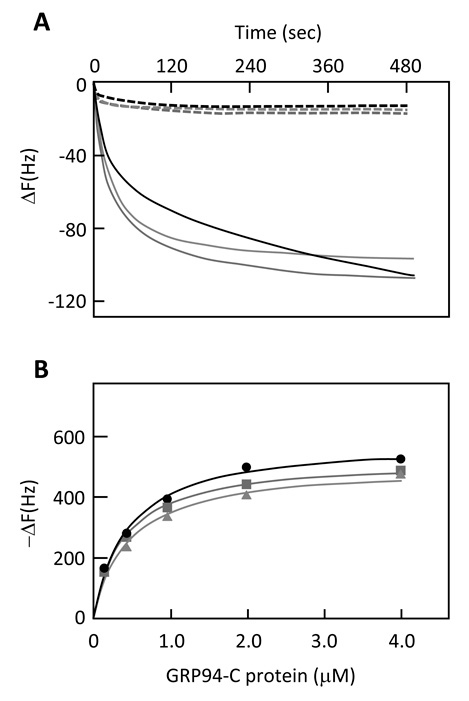
Fig 5.
QCM analysis. (A) Typical time-course plots of the Frequency changes (ΔF) after addition of GRP94-C protein (solid line) or GST protein (dot-line) to peptide No.6 (black), No.58 (red) or No.72(blue)-immobilized in cells in 27-MHz quartz plates. Proteins were added to the cells at the concentration of 250 nmol/L. Data are from one representative experiment out of at least three performed. (B) Plots of ΔF at equilibrium against the concentration of GRP94-C protein. DF responding to the addition of GRP94 to peptide No.6, No.58 and No.72-immobilized in plated cells are plotted with black, red, and blue lines, respectively.
Low bioavailability and nonspecific toxicity to healthy tissues present major obstacles in the development of new therapeutic anticancer drugs. Discovery of functional ligand-receptor systems is one of the critical steps toward the development of an efficient cancer therapy. Recently, several studies have suggested that proteins selectively expressed on the surface of cancer cells are potentially useful targets for the development of novel anticancer therapies[5, 17].
Previous studies have shown that the GRP94 chaperon protein is expressed on the cell surface in some cancer cells but not in non-transformed cells[12]. Characteristics of cell surface expression of GRP94 have however received much less attention, since it is widely considered to be a chaperon in immune signaling and peptide presentation for antigen representing cells[18]. In the present study, we focused on cell surface expression of GRP94 as a target for a peptide-drug conjugate. As a first step, we screened GRP94 protein-binding peptides using a phage display technique. The C-terminal domain of GRP94 protein was chosen to avoid non-specific, physiological peptide binding of the N-terminal domain[14]. Initially, 16 peptides were selected from the phage display-derived pool of candidate peptides and divided into four groups based on their amino acid similarity (Fig. 1). Threonine, proline and leucine amino acid residues were frequently found in the peptide sequences for each of the four groups and these may play a role in GRP94 protein binding.
Three peptides (peptide Nos. 6, 58 and 72) out of the above-mentioned 16 candidates were selected based on their higher absorbance at 405 nm using multiple ELISAs and gave values within a similar range (absorbance reading 1.3-1.6 at 405 nm). QCM analysis revealed a high affinity of these peptides to GRP94-C protein. Kd values for the binding of all three peptides to GRP94 were within a similar range of 0.39-0.46 uM, which also corresponded well to the similar range of absorbance levels obtained using ELISA (Fig. 4,Fig. 5). The GRP94-C protein did not bind to either the random peptide in ELISA assay or the GST protein in QCM analysis, which further confirmed the specificity of our peptides towards the C-terminal of the GRP94 protein.
In summary, we have reported for the first time the discovery of synthetic peptide ligands that bind to the C-terminal domain of the GRP94 protein with high affinity. Our results suggest that these novel peptide sequences could potentially be used as a part of a peptide-conjugated drug system, designed to selectively target cancer cells with surface expression of GRP94.
This work was supported in part by Grants-in-Aid for Scientific Research through the Industrial Technology Research Grant Program in ‘04 from the New Energy and Industrial Technology Development Organization, the Smoking Research Foundation, the Tokyu Foundation for a Better Environment, the Kieikai Research Foundation, and the Japan Society for the Promotion of Science. M. Zahed was a doctoral student supported financially by a MEXT Honors Scholarship for International Students.
【目的】
癌に特異的に発現する細胞表面タンパク質に結合するペプチドを利用した,癌細胞特異的薬物輸送法が考案されている。ストレスタンパク質GRP94は,細胞の造腫瘍化に伴って,細胞表面の発現量が増加する。そこで,本研究では,ファージディスプレイ法を用い,GRP94タンパク質のカルボキシ末端領域(GRP94-C) へ結合する低分子ペプチドの探索を行った。
【方法】
ランダムな7アミノ酸配列の両端をシステイン残基のジスルフィド結合で環状化したペプチド配列を持つファージディスプレイライブラリーを使用した。組換型GRP94-Cタンパク質に結合するペプチド配列の濃縮を3回繰り返した後,濃縮された任意の80クローンのアミノ酸配列を調べた。コンセンサス配列を含むクローンについて結合アッセイ,QCM解析を行った。
【結果】
3つのペプチド配列(No.6; CTLMSDPAC, No. 58; CSLVPIPAC, No. 72; CQTKPLFQC) を見出した。QCM法により3ペプチド配列のGRP94-Cタンパク質に対する結合定数を測定したところ,それぞれ,0.45,0.46,0.39 uMであった。
【考察】
本研究により,GRP94-Cタンパクへ高親和性で結合するペプチド配列を見出すことが出来た。これらペプチドを利用して,細胞表面にGRP94を高発現している癌細胞特異的な薬物輸送への応用利用が期待される。
ザヘッド ムハメド1),鈴木敏和1,2),喜多和子1),菅谷 茂1),鈴木信夫1) :ファージディスプレイ法を用いたGRP94タンパク質カルボキシ末端側領域に結合するペプチドの探索.
Tel. 043-290-3118. Fax. 043-290-3118. E-mail: t-suzuki@wayo.ac-jp
2010年11月18日受付,2010年12月20日受理.
