




Chiba Medical J. 94E:45-50,2018
doi:10.20776/S03035476-94E-5-P45
[ Original Article ]
Daisuke Ishii1,3)*, Daisuke Matsuzawa1,2), Shingo Matsuda1,4,5),
Haruna Tomizawa-Shinohara1), Chihiro Sutoh1),
Yutaka Kohno3) and Eiji Shimizu1,2)
1 ) Department of Cognitive Behavioral Physiology, Chiba University Graduate School of Medicine, Chiba 260-8670 .
2 ) Research Center for Child Mental Development, Chiba University Graduate School of Medicine, Chiba 260-8670 .
3 ) Center for Medical Sciences, Ibaraki Prefectural University of Health Sciences, Ibaraki 300-0394 .
4 ) Department of Pharmacotherapeutics, Showa Pharmaceutical University, Tokyo 194-8543 .
5 ) Department of Ultrastructural Research, National Institute of Neuroscience, National Center of Neurology and Psychiatry, Tokyo 187-8502 .
(Received March 12, 2018, Accepted May 15, 2018)
Fear memory is more stable and stronger than extinction memory itself, and is stored for a longer period of time. However, it is unclear whether fear memory can last for a lifetime. We examined the ability to store contextual and cued fear memory in 90-week-old mice that had received fear conditioning and cued fear extinction at 12 weeks of age. Our major findings were as follows: 1) 78 weeks after the fear conditioning, contextual fear memory was erased. 2) 78 weeks after the cued fear extinction, recovery of fear was not observed. These results suggested that the contextual and cued fear memory could not last for a lifetime. Our results that the memories of stressful events did not last for a lifetime could be helpful for clarifying the mechanisms of memory storage.
fear memory, fear extinction, spontaneous recovery of fear, fear renewal, aging
The most widely acknowledged psychological change with age is the decline in memory ability. Alzheimer’s disease, known as one of memory impairment, causes memory loss [1]. On the other hand, there is a report that people with Alzheimer’s disease remembered the episode of the earthquake experienced two months ago[2]. Moreover, it has been reported that fear memories were preserved without erasing for at least 480 days in rats [3]. These studies suggest that fear memory is easy to acquire and stable for a long time.
Fear is an important emotion for risk assessment, but an excessive fear causes anxiety disorders such as posttraumatic stress disorder(PTSD). Patients with PTSD, contrary to Alzheimer’s disease that cause memory loss, have difficulty in erasing the acquired fear memory. To investigate the difference in characteristics depending on the memory types could be helpful for clarifying the pathology of these memory-associated diseases.
The paradigm of fear conditioning and extinction in rodents has been considered a valuable animal model for studying the memory process of fear [4,5]. Fear conditioning forms the associative learning between a conditioned stimulus(CS)such as a tone(hippocampus-independent)or context(hippocampusdependent)and an aversive unconditioned stimulus(US) such as an electric footshock[6]. Fear extinction refers to the repeated presentations of the CS in the absence of the US previously paired. It has been hypothesized that fear extinction forms a new memory of safety that inhibits fear expression but does not erase the original fear memory[7]. Accordingly, fear that is reduced by extinction can be recovered spontaneously after the passage of time (spontaneous recovery); moreover, it can be renewed by the presentations of the CS alone outside of the extinction context(renewal)[8]. These studies suggest that the process of fear memory is different from that of extinction memory. Furthermore, fear memory is more stable and stronger than extinction memory itself.
Given this background, we investigate whether aged mice that had received fear conditioning in their young age could retrieve their contextual fear memory (hippocampus-dependent). Secondly, we examined whether the recoveries of a cued fear memory(hippocampus-independent)extinguished by cued fear extinction would be observed after the passage of a long period of time since the cued fear extinction.
C57BL/6J male mice were housed 5 per cage from the age of 3 to 90 weeks under a 12-h light/dark cycle(lights on at 07 : 00 hours) and controlled temperature(23±1℃). The animals were provided food and water ad libitum. Each behavioral testing was conducted between 09 : 00 and 12 : 00 hours. Mice were used at 12 and 90 weeks of age. The research and animal care were carried out according to the Guide for Animal Experimentation of the Chiba University Graduate School of Medicine(25-134).
To examine whether acquired contextual fear memory would be stored at from 12 weeks to 90 weeks of age, and whether the cued fear memory that was reduced by cued fear extinction would recover, we trained mice using a mixed fear conditioning, cued fear extinction, contextual test, cued test(spontaneous recovery) and renewal test paradigms(Fig. 1).
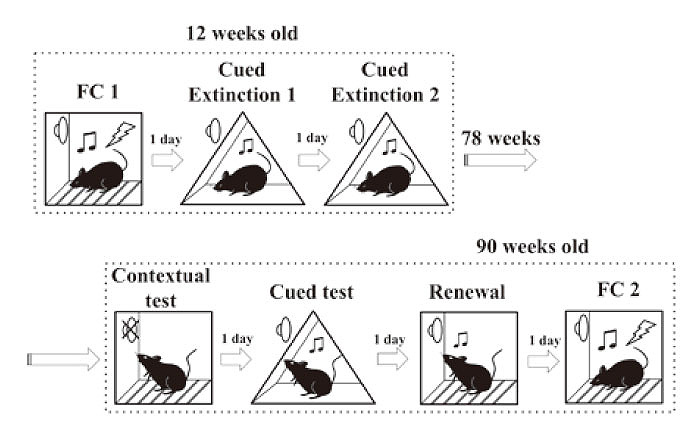
Fig. 1
Schematic representations of the experimental schedule. Fear conditioning was performed in a square box(context A) at 12 weeks of age. 24 hours later, cued fear extinction 1 was performed in a triangular box(context B). 24 hours after extinction 1, extinction 2 was performed in context B. 78 weeks later, a contextual memory test was performed in context A that had been used for fear conditioning at 12 weeks of age. 24 hours later, a cued memory test was performed in context B that had been used for extinction sessions at 12 weeks of age. 24 hours after the cued memory test, a renewal test was performed in context A. 24 hours later, fear conditioning trial 2 was performed in context A in mice 90 weeks of age.
As described in detail previously[9], all mice were received 3 pairings(90-150 sec interstimulus interval) of a conditioned stimulus(CS; 30 sec, 80 dB, 2.8 kHz tone) and an unconditioned stimulus(US; 2 sec, 0.75 mA scrambled footshock). The US was presented during the last 2 sec of the CS. After the final CS-US pairing, mice were returned to the home cage.
On day 2 and 3, extinction sessions divided into 2 sessions(extinction 1 and 2) were performed in a novel context(extinction chamber). In extinction 1, all mice received 20 presentations of the tone alone(interstimulus interval: 5 sec) after an initial 180 sec pre-exploration period. Twenty-four hours after extinction 1, all mice received extinction 2, which consisted of 40 tones(interstimulus interval: 5 sec) after an initial 180 sec pre-exploration period.
Seventy-eight weeks after extinction 2, all mice were returned to the conditioned chamber to test their contextual fear memory, and remained in this setting for 10 minutes.
Twenty-four hours after the contextual test, all mice were returned to extinction chamber to test their spontaneous recovery of fear. After an initial 100 sec pre-exploration period, all mice received 4 presentations of the tone alone (interstimulus interval: 50 sec). Twenty-four hours after the spontaneous recovery test, all mice were placed in the conditioned chamber and presented with 4 CS to measure their level of fear renewal.
Twenty-four hours after the renewal test, phase 2 of the fear conditioning was performed by repeating the behavioral procedures used for the first phase of fear conditioning.
As described in detail previously[9], the activity of mice was monitored by FreezeFrame software(Actimetrics Software, Wilmette, IL). Freezing(no visible movement except respiration) was scored every 5 sec and covered to a percentage[ (freezing observations/total observations)×100]. The data from the fear extinction experiments 1 and 2, and the comparisons of the percentages of freezing mice among the first 4 trials of the extinction 1, contextual test, cued test and renewal test were analyzed using one-way repeated measures of analysis of variance(ANOVA). In the analysis of extinction phases, we pooled the trials into 4(extinction 1) and 8(extinction 2) bins in order to avoid potential type 1 errors for multiple comparisons. On the other hand, the data from the comparison of the fear conditioning phases 1 and 2 were analyzed using two-way repeated measures of ANOVA. Bonferroni ’s correction was used for post hoc comparisons when ANOVA revealed statistically significant differences. Statistical significance was set at p<0.05 in all the analyses. All analyses were performed with the software package SPSS 12.0 for Windows(SPSS, Chicago, IL). Data are shown as the mean±SEM for all the results.
In the fear extinction phase 1, one-way repeated ANOVA showed no significant main effect of bins(F(3, 12)=2.454, p>0.05)(Fig. 2).
In the fear extinction phase 2, one-way repeated ANOVA showed a significant main effect of bins(F(7, 28)=4.434, p<0.01), (Fig. 2). Post hoc comparison revealed that the % freezing value at the 1st bin was significantly higher than those at the 4th, 5th, 6th and 8th bins(p<0.05, p<0.01, p<0.05, and p<0.01, respectively)(Fig. 2).
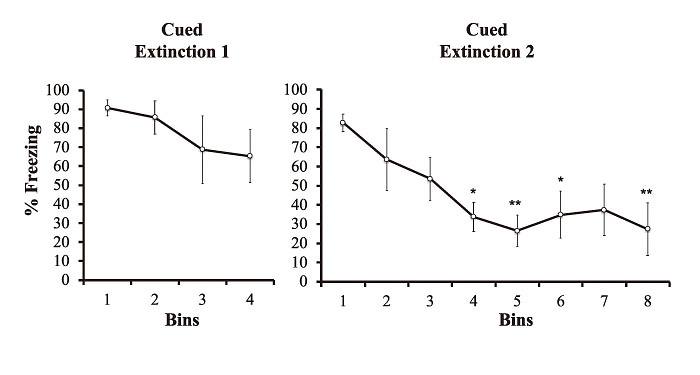
Fig. 2
Cued fear extinction 1 and 2. The % freezing value to the tone during extinction phases 1 and 2 at 12 weeks of age(n=5). The data were pooled into 4(extinction 1) and 8(extinction 2) bins. A bin is the average of 5 trials of extinction. All data points are the means±SEM. *p<0.05 and **p<0.01 indicate a statistically significant difference from the first bin(the first 5 trials) of extinction phase 2.
In the first 4 trials of the extinction 1, contextual test, cued test and renewal test, one-way repeated ANOVA showed a significant main effect of test(F(3, 12)= 182.565, p<0.01). Post hoc comparison indicated that the % freezing value during the first 4 trials of the extinction 1 at 12 weeks of age was significantly higher than the % freezing values at the contextual test, cued test and renewal test at 90 weeks of age(p<0.01, p<0.01, and p<0.01, respectively)(Fig. 3).
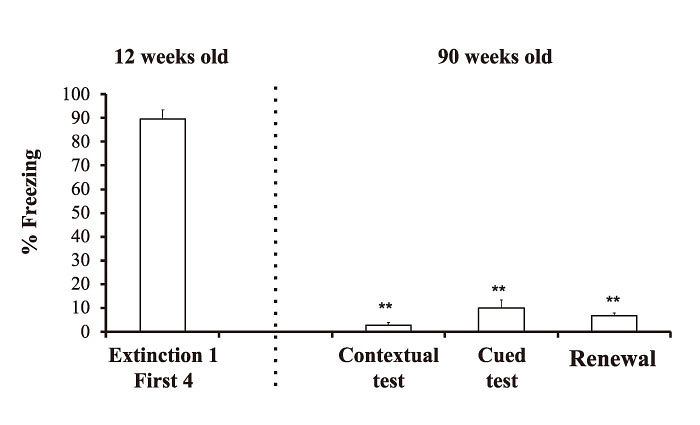
Fig. 3
The % freezing values during the first four trials of extinction 1 (First 4), contextual memory test(contextual test), cued memory test(cued test) and renewal test. Fear extinction sessions were performed at 12 weeks of age. A contextual test, cued test and renewal test were performed at 90 weeks of age. All data points are the means ± SEM. **p<0.01 indicates a statistically significant difference from the first 4 trials of extinction phase 1.
In the fear conditioning phases 1 and 2, two-way ANOVA with repeated measures showed a significant main effect of trial (F(2, 16)=48.234, p<0.01), group (F(1, 8)=25.14, p<0.01) and interaction between group and trial(F(2, 16)=6.209, p<0.05). Post hoc comparison revealed that the % freezing value in 90-week-old mice was significantly lower than that in 12-week-old mice in trials 2 and 3(p<0.01, p<0.01, respectively)(Fig. 4). Moreover, we found that %freezing value during the trial 2 and 3 in 12 weeks old group significantly higher than that at the 1st trial(p<0.01, p<0.01, respectively). The %freezing value during the trial 3 in 90 weeks old group significantly higher than that at the 1st trial(p<0.01).
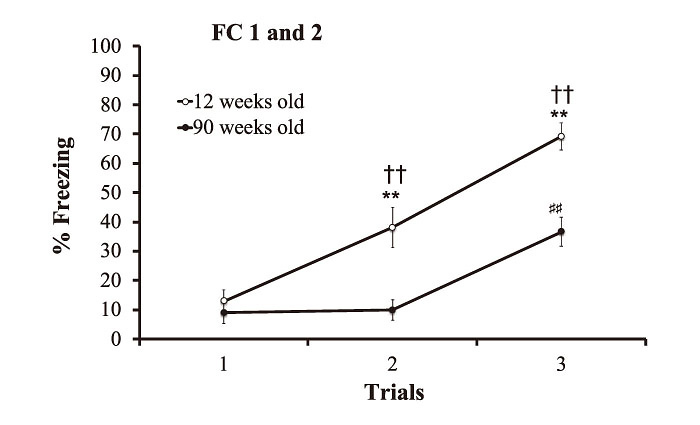
Fig. 4
The % freezing values to the tone during fear conditioning phase 1 at 12 weeks of age and fear conditioning phase 2 at 90 weeks of age. All data points are the means±SEM. **/##p<0.01 indicates a statistically significant difference from the first trial of fear conditioning in 12 weeks old group/90 weeks old group. ††p<0.01 indicates a statistically significant difference between the 12-week-old and 90-week-old mice.
The major findings of our study were as follows: 1) seventy-eight weeks after fear conditioning, contextual fear memory was erased; 2) seventy-eight weeks after cued fear extinction, no recovery of the cued fear memory was observed.
Acquired fear memory is known to be recalled 480 days after fear conditioning in rats[3]. In our study using mice, contextual fear memory acquired at 12 weeks of age, which was not extinguished by contextual fear extinction, could not be recalled at 90 weeks of age (546 days after fear conditioning). There is a possibility that difference in species caused these conflicting results. In addition, spontaneous recovery and fear renewal were not observed 78 weeks after fear conditioning. There are reports that spontaneous recovery is observed 1 month after fear conditioning and fear renewal is observed 2 weeks after fear conditioning[10,11]. These studies suggest that cued fear memory could be erased by the passage of a long period of time after fear conditioning.
It has been reported that animals in which a previously conditioned fear memory had been erased showed a low rate of reacquisition of fear memory, relative to a group being conditioned for the first time[11]. Our current study revealed that the rate of fear memory acquisition at 90 weeks of age was lower than that at 12 weeks of age, suggesting that cued and contextual fear memories are completely erased after 78 weeks from fear conditioning.
Although we observed that aged mice succeeded in the acquisition of cued fear memory, it should be noted that C57BL/6J mice begin to have frequencyspecific hearing loss from about 10 weeks of age. Such hearing loss increases in severity, spreading to lower frequencies as the age advances over the next 12 to 15 months[12]. In another previous study, however, aged C57BL/6 mice were shown to have intact acquisition of cued and contextual fear memories [13]. In addition, we confirmed that aged mice exhibited significantly high freezing behavior during a cued fear memory test performed after the second phase of fear conditioning(data not shown). Although a possibility that 90-weekold mice in our study experienced partial hearing loss cannot be ruled out, we prefer the interpretation that the original fear memory was completely erased by the passage of a long period of time after the fear conditioning, because these aged mice could reacquire and recall a reconditioned fear memory to tone. Furthermore, it has been reported that the motor activity of C57BL/6J male mice decreases with age, and more time is spent in inactivity[14]. In our study, decreased motor activity of aged mice did not affect the behavioral results, because aged mice exhibited a low freezing level during cued and renewal tests. We did not observe the unusual high freezing behavior in aged mice during fear conditioning phase 2.
A previous study has shown that acquired contextual fear memories were initially stored in the hippocampus, but that these memories progressively transferred from the hippocampus to an extra-hippocampal region, such as the neocortex[15]. This previous report and our own present study suggest the possibility that contextual fear memory may gradually weaken if it is never recalled after its transition to the neocortex.
Numerous studies have reported the mechanisms of fear memory formation, fear extinction and recovery of fear memory using C57BL/6J male mice[9,16-22]. With respect to fear memory, studies on aging using C57BL/6J male mice should take into consideration our present finding that this strain could not retain contextual and cued fear memory for 78 weeks. Our study suggests that the acquired fear memory storage could be vanished with aging, although it should be examined whether or not those memory erasure would depend on memory types.
DI, DM, SM, HT, CS, YK and ES conceived of the presented idea. DI, DM, SM and HT designed the behavioral experiments. DI, SM and HT carried out the experiments. SM, HT and CS analyzed all data. DI, DM, SM, YK and ES interpreted results. DI and DM drafted the manuscript. All authors approved the final version of the submitted manuscript.
This work was supported by JSPS KAKENHI Grant Number JP 26461711.
The authors disclose no conflicts of interest.
Address correspondence to Dr. Daisuke Ishii.
Department of Cognitive Behavioral Physiology, Chiba University Graduate School of Medicine, 1-8-1, Inohana, Chuou-ku, Chiba 260-8670, Japan.
Phone: +81-29-840-2315. Fax: +81-43-226-2028.
E-mail:d-ishii@umin.ac.jp
