




Chiba Medical J. 96E:1-4, 2020
doi:10.20776/S03035476-96E-1-P1
[ Chiba Medical Society Award Review ]
Shigeru Tanaka
Department of Allergy and Clinical Immunology, Graduate School of Medicine, Chiba University, Chiba 260-8670.
(Received November 4, 2019, Accepted November 10, 2019, Published February 10, 2020.)
Regulatory T (T reg) cells play critical roles in maintaining immune homeostasis, and the development of this unique T cell subset is dependent on the master transcription factor, Foxp3. The transcription of Foxp3 is tightly regulated by several regulatory elements. While there is accumulating evidence that multiple transcription factors promote Foxp3 expression at steady-state conditions, little is known about how Foxp3 expression is controlled under inflammatory conditions. Although the expression of Foxp3 is sufficient to induce the suppressor function in murine CD4+ T cells, Foxp3- binding proteins are required for the appropriate T reg suppressor function. Thus, the precise understanding of Foxp3-containing transcription complex is important for optimizing T reg cell-based therapy for autoimmune diseases. In this review, the molecular mechanisms underlying the regulation of Foxp3 expression, especially under inflammatory conditions, are summarized. Further, Foxp3 partner proteins and T reg cell function are also discussed.
regulatory T cells, Foxp3, Sox12, KAP1
Regulatory T (T reg) cells are indispensable for immune tolerance and suppress excessive inflammation. In 1995, Dr. Sakaguchi’s group showed that CD4+ CD25+ T cells have immunosuppressive function in mice as antibody-mediated depletion of this population resulted in the development of autoimmune diseases [1]. A few years later, his and Dr. Rudensky’s group independently reported that Foxp3 is the master transcription factor of CD4+ CD25+ T reg cells [2,3]. Of note, mutations in FOXP3 gene in humans result in the development of immune dysregulation, polyendocrinopathy, enteropathy, X-linked (IPEX) syndrome; patients with IPEX syndrome acquire multiple autoimmune diseases, indicating the crucial role of Foxp3 in regulating immune responses. Over the last decade, there has been a tremendous interest in the field of T reg-related research and researchers have shown the importance of T reg cells not only in autoimmune diseases, but also in cancer[4], metabolic disorders[5], and transplantation[6].
The expression of Foxp3 is a crucial step toward the development of T reg cells because the ectopic expression of Foxp3 is sufficient to induce suppressive capacity in conventional CD4+ T cells[2]. The transcription of Foxp3 is regulated by the promoter and three enhancers located within the conserved non-coding sequences named CNS1, 2, and 3. Zheng et al. generated three knockout mouse lines lacking Foxp3 CNS1, 2 or 3, and the analyses of these CNS knockout mice revealed that CNS1, 2, and 3 are indispensable for T reg cell differentiation in the peripheral tissues, T reg stability, and T reg cell differentiation initiation, respectively[7]. Recently, a new Foxp3 enhancer region named CNS0 which works as a super enhancer has been defined[8]. As mentioned above, T reg cell differentiation in the periphery is mainly controlled by CNS1 and it is shown that Smad3 and NFAT are recruited to CNS1 upon TGF- β and T cell receptor (TCR) stimulation, respectively [9]. All-trans-retinoic acid (ATRA) , a vitamin A metabolite, also induces T reg cell differentiation by boosting the Smad3 signaling[10]and suppresses Th17 cell differentiation. In addition, AP-1 activates CNS1 and promotes T reg cell differentiation in the periphery [11]. However, little is known about how Foxp3 expression is regulated under inflammatory conditions such as colitis, because most of these studies were conducted under steady-state conditions.
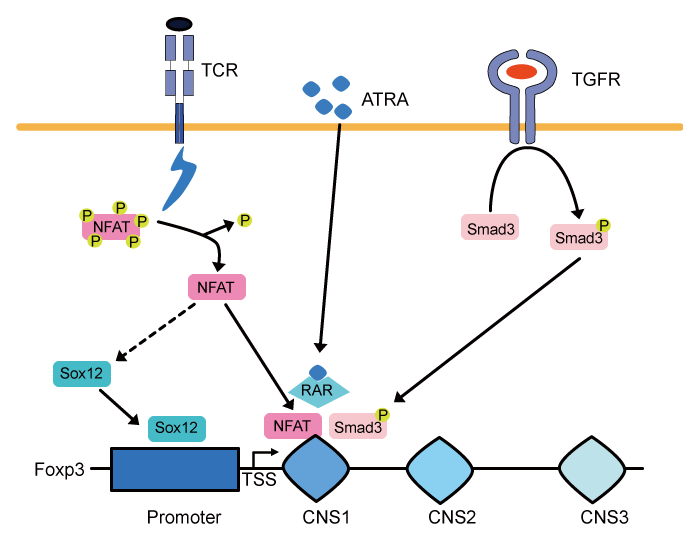
Fig.1 Signals up-regulating Foxp3 transcription under inflammatory conditions.
NFAT is activated and recruited to the Foxp3 CNS1 enhancer, when conventional CD4+ T cells recognize their own antigen. In addition, activated NFAT also promotes Sox12 expression and this leads to further activation of the Foxp3 promoter region. ATRA-induced retinoic acid receptor (RAR) activation and TGF-β-mediated Smad3 phosphorylation also support the expression of Foxp3 in the peripheral tissue under inflammatory conditions.
TSS: transcription start site.
The author and colleagues have performed RNAseq analyses of T reg cells recovered from mice with dextran sodium sulfate colitis to find novel T reg cellrelated transcription factor (s) upregulated during colitis[12]. Among 56 genes which are differentially expressed in T reg cells of colitic mice, we focused on a transcription factor Sox12, a member of SoxC family which is composed of Sox4, Sox11, and Sox12. It has been shown that SoxC is critical for the development of the nerve system[13]. In addition to the developmental function, accumulating evidence indicates their roles in the immune system. For example, Kuwahara et al. demonstrated that Sox4 negatively regulates Th2 differentiation by interfering with Gata-3 function[14]. On the other hand, Yoshitomi et al. have shown that Sox4 facilitates the development of CXCL13-producing helper T cells in autoimmune diseases in humans[15]. These results suggest the crucial roles of SoxC family members in the acquired immunity.
We first analyzed the expression of SoxC family in CD4+ T cells. In vitro, the expression of Sox12 was induced, whereas the expression of Sox4 was dramatically reduced by TCR stimulation in both conventional CD4+ T cells and T reg cells. In contrast, Sox11 was not expressed in CD4+ T cells even after TCR stimulation. These findings suggest distinct roles of Sox4 and Sox12 in CD4+ T cells.
We also found that while Sox12 knockout mice did not develop autoimmune diseases spontaneously, Sox12-deficient CD4+ T cells were colitogenic in the T cell transfer model of colitis. Importantly, when Sox12- deficient naïve T cells and wild type (WT) naïve Tcells were co-transferred into lymphopenic RAG2 knockout mice, Foxp3 positive cells in Sox12-deficient T cells were less frequent as compared to those in WT T cells, indicating the role of Sox12 in the differentiation of T reg cells in the peripheral tissue during colitis. Consistently, over-expression of Sox12 efficiently promoted Foxp3 expression in naïve CD4+ T cells, and Sox12-mediated Foxp3 expression was still observed even in the presence of antibody against TGF-β or IL- 2, both of which are important for the induction of T reg cells. In addition, Sox12-expressing cells showed the suppression function both in vitro and in vivo. Chromatin immunoprecipitation (ChIP) analyses and reporter assays revealed the direct effect of Sox12 in promoting Foxp3 induction by binding to the Foxp3 promoter. These results suggest that TCR-mediated up-regulation of Sox12 during colitis favors T reg cell differentiation in the periphery and protects mice from excessive inflammation.
Although the forced expression of Foxp3 is sufficient to induce suppressive T reg cells, other transcription factors that physically interact with Foxp3 are required for optimal T reg function. Rudensky and his colleagues carried out a comprehensive analysis of Foxp3-binding proteins and showed that more than 300 proteins could potentially associate with Foxp3 [16]. One of such partner proteins is Gata-3. A part of thymus-derived T reg cells express Gata-3; T regspecific Gata-3-deficient mice spontaneously develop autoimmune diseases such as colitis and dermatitis v. In addition to Gata-3, Eos, HDACs, RUNX1, NFAT, and NF-κB are also reported as partners of Foxp3 [17]. Intriguingly, T reg cells that lack any of these Foxp3-binding partners fail to exhibit normal suppressor function. A recent study has shown that Foxp3 serves as an activator when it is complexed with RELA, IKZF2, and KAT5. In contrast, Foxp3 functions as a repressor when complexed with EZH2, YY1, and IKZF3[18]. These findings lead to speculation that the transcription program controlled by Foxp3 is tightly regulated and each of the Foxp3-binding partners is required for the optimal transcriptional landscape of T reg cells.
Dr. Ziegler and his colleagues reported a novel FOXP3-interacting protein, FIK (FOXP3-interacting KRAB domain-containing protein) in human T reg cells [19]. FIK is a novel splicing isoform of ZFP90 and acts as a bridging molecule that connects FOXP3 with KAP1 (also known as TIF-1β/ TRIM28) ; the knock-down of FIK or KAP1 results in decreased suppression capacity in human T reg cells.
KAP1 is a member of the tripartite motif proteins and was originally reported as a chromatin remodeling factor. KAP1 is critical for heterochromatin structure and is highly related to suppressive H3K9me3 mark. For example, KAP1-mediated deposition of H3K9me3 is an important mechanism to silence endogenous retroviral elements[2]. However, considering the interaction between FOXP3 and FIK/KAP1 in human T reg cells and based on a previous report showing antiinflammatory effects on conventional T cells[21], we hypothesized that KAP1 has different roles in T reg cells.
We generated T reg-specific KAP1-deficient mice (T regΔKAP1) to address the mechanisms by which KAP1-Foxp3 complex regulates T reg development and function[22]. Interestingly, adolescent T regΔKAP1 mice showed a lymphoproliferative disease and lung inflammation although the development of T reg cells in the thymus was not disturbed. T regΔKAP1 mice also exhibited elevated anti-nuclear antibody, suggesting that T regΔKAP1 mice develop global loss of tolerance. Consistent with the phenotype observed in T regΔKAP1 mice, KAP1-deficient T reg cells had diminished suppression function both in vitro and in vivo. ChIPseq analyses revealed that almost half of genomic loci occupied with Foxp3 were also bound by KAP1. Intriguingly, most of these genes regulated by KAP1- Foxp3 complex were immune-related genes, suggesting the critical role of KAP1 in T reg cells. For example, Prdm1 (encodes Blimp-1) is co-occupied with KAP1 and Foxp3, and the expression of Blimp-1 is decreased in KAP1-deficient T reg cells. As a previous report showed important roles of Blimp-1 in the production of immuno-suppressive cytokine IL-10[23], we analyzed IL-10 expression in T reg cells in T regΔKAP1 mice[22]. Although IL-10 production in spleen was comparable between the control and T regΔKAP1 mice, IL-10-producing T reg cells were reduced in the lung of T regΔKAP1 mice where the spontaneous inflammation was observed. These results suggest that KAP1-Foxp3 complex governs T reg suppressor function through the expression of T reg-associated genes (e.g. Prdm1)
Although the prognosis of autoimmune diseases is gradually improving, the patients must continue to take immune-suppressive agents including glucocorticoids for life; the long-term use of non-specific immunesuppressive drugs can lead to adverse events such as serious infections. Researchers are now developing T reg cell-based therapy because this new therapeutic strategy is expected to have less toxicity. However, we need to improve this new therapy, as the efficacy is still insufficient. Further progress in T reg research field is expected to overcome the autoimmune diseases in the future.
I thank Drs. Hiroshi Nakajima and Akira Suto for their help and critical reading of this manuscript and Dr. Steven Ziegler for kind support.
The author declares no competing interests.
Address correspondence to Dr. Shigeru Tanaka.
Department of Allergy and Clinical Immunology, Graduate
School of Medicine, Chiba University, 1-8-1, Inohana, Chuouku,
Chiba 260-8670, Japan.
Phone: 81-43-226-2198. Fax: +81-43-226-2199.
E-mail:stanaka@chiba-u.jp
