




Chiba Medical J. 97E:39-48, 2021
doi:10.20776/S03035476-97E-4-P39
〔 Original Article 〕
Manabu Arai1) , Masahiro Sakakibara1) , Takeshi Nagashima2)
and Masayuki Otsuka2)
1) Departments of Breast Surgery, Toho University Sakura Medical Center, Sakura 285-8741.
2) Departments of General Surgery, Chiba University Graduate School of Medicine, Chiba 260-8670.
(Received November 6, 2019, Accepted April 12, 2021, Published June 10, 2021.)
【Background】Cancer and inflammation have long been considered two sides of the same coin. With the identification of a subset of CD4 (+) T cells that produce interleukin (IL) -17, called Th17 cells, the relationship between this subset and tumor-specific inflammation in the tumor microenvironment has been brought back into the spotlight. The purpose of this study was to determine relationships between Th17 cells and both tumor progression and lymph node invasion, and to evaluate cytokine balance in the tumor microenvironment with tumor progression in patients with breast cancer.
【Methods】Tumor tissues and sentinel lymph nodes were collected from 50 breast cancer patients who had been treated at our hospital. Expressions of retinoid orphan nuclear receptor C (RORC) , IL-17, IL-21, IL-6, tumor growth factor (TGF) -β, IL-2 and forkhead box P3 (Foxp3) were examined by measuring their mRNA levels with real-time RT-PCR, and correlations of tumor progression to Th17 cells and surrounding cytokines were investigated.
【Results】mRNA levels of the RORC gene, which serves as the master gene for Th17, were elevated with increasing clinical stage of breast cancer in both the main tumor and sentinel lymph nodes. In the tumor microenvironment, IL-17 mRNA increased in parallel with RORC expression, while IL-2 mRNA did not increase. TGF-β and IL-6 mRNA levels were elevated with tumor progression, closely correlating with RORC mRNA levels.
【Conclusions】Biomarker expression for Th17 cells increased with tumor progression in the tumor microenvironment of human breast cancer. Th17 cells may play a key role in the malignant cycle of tumor progression and tumor-specific inflammation maintenance.
breast cancer, Th17 cells, immunity, inflammation
Breast cancer is the most common malignancy in women. With revelations about the microenvironment in cancer[1-3], interactions between cancer cells and stroma cells in a cytokine environment based on inflammation (involving tumor growth factor (TGF) -β and interleukin (IL) -6) have been brought back into the spotlight [4,5]. A previous study showed that increased levels of infiltrating T cells in tumor represent a good prognostic factor [6-8]. However, the interaction between T cells, which control inflammation in normal immunity, and cancer cells was unclear. A subset of CD4 (+) T cells, called T helper (Th) 17 cells, has been revealed [9- 11]. In animal models, Th17 cells are generated in the presence of TGF-β and IL-6, expand under the influence of IL-23 and produce IL-17[12-15]. Retinoid orphan nuclear receptor gamma-t, for which the human homolog is retinoid orphan nuclear receptor C (RORC) , has been considered the master transcriptional regulator of Th17 cell differntiation[16]. A previous study reported that RORC opposes the action of forkhead box P3 (Foxp3) , which is known as the pivotal transcription factor for regulatory T (Treg) cell maturation[17]. Th17 cells are associated with responses against certain infections, are implicated in the development of autoimmunity and play a key role in chronic inflammatory diseases[18- 22]. Previous studies have shown that IL-17 and IL-23 are associated with tumor progression and angiogenesis [23,24]. Th17 cells have been implicated in various tumors[25-28]. Kryczek et al. reported the presence of Th17 cells in tumor and draining lymph nodes in human and mouse tumors[25]. Previous results have suggested the potential of Th17 cells to play a central role in progression of the tumor microenvironment based on inflammation[23].
Several studies have demonstrated the presence of Th17 cells in human breast tumors[29-35]. The purpose of this study was to determine whether Th17 cells increase with tumor progression and lymph node invasion, and to evaluate the cytokine balance in the tumor microenvironment among patients with breast cancer.
Patients and tissue samples
Tumor tissues and sentinel lymph nodes (SLNs) were collected from 50 breast cancer patients (mean age, 52 years; range, 32-68 years) who had been treated in the Department of General Surgery at Chiba University Graduate School of Medicine (Chiba, Japan) between January 2006 and December 2007. We selected 50 patients at stage I (n=20) , stage IIA (n=10) , stage IIB (n=10) or stage IIIA (n=10) of the Union for International Cancer Control (UICC) tumor/node/metastasis (TNM) classification in order of operation date. SLNs and tumor were collected during surgery. None of the patients had received radiotherapy, chemotherapy, or other medical interventions prior to the study. Patient characteristics are summarized in Table 1.
Real-time RT-PCR
From 50 patients with breast cancer, tumor tissue and one terminal section of the SLN that had not been used for pathological diagnosis were frozen immediately. Total RNA was extracted from these tumor and SLN tissues using an RNeasy Mini Kit (QIAGEN, Hilden, Germany) . The prepared total RNA served as the template for first-strand complementary DNA (cDNA) synthesis using Ready-To-GoTM You-Prime First-Strand Beads (Amersham Pharmacia Biotech, Piscataway, NJ, USA) . The cDNA product was analyzed at a final Mg concentration of 3 mM in a LightCycler with software (version 3.5; Roche Molecular, Mannheim, Germany) using a FastStart DNA Master SYBR Green I Kit (Roche Diagnostics, Mannheim, Germany) . PCR was performed with the following primer sets: RORC, forward 5’- GGA CCA AGA CAT CCT CAA GA -3’, reverse 5’- GTC TCT GAG CTT GAT CCA TCT -3’; IL-17, forward 5’- TGT GCC AGG AGG TAG TAT GA -3’, reverse 5’- ACA CTG GGC CTG TAC AAC TT -3’; IL-6, forward 5’- GGA CCA AGA CAT CCT CAA GA- 3’, reverse 5’- ACA CTG GGC CTG TAC AAC T- 3’; TGF-β, forward 5’- TGA TGT CAC CGG AGT TGT G -3’, reverse 5’-TGA ACC CGT TGA TGT CCA -3’; IL-21, forward 5’- AGG TCA AGA TCG CCA CAT -3’, reverse 5’- GAC TTT AGT TGG GCC TTC TG -3’; Foxp3, forward 5’- AGG ACA GCA CCC TTT CGC CT -3’, reverse 5’- GAC ATT GTG CCC TGC CCT TC -3’; IL-2, forward 5’- GAA GAA CTCAAACCTCTG GA -3’, reverse 5’- GTC TCA TCA GCA TAT TCA CAC -3’; and glyceraldehyde-3- phosphate dehydrogenase (GAPDH) , forward 5’- GTC CTT GGC GGG TAA TTC TA -3’, reverse 5’- GCT GAA GTC TGG CTT CTT GG -3’. For each primer, a standard curve was developed. PCR conditions for RORC, IL-17, IL-21, IL-6, TGF-β, IL-2, Foxp3 and GAPDH were as follows: 95 ℃ for 10 min, then 40 cycles at 95 ℃ for 15 s, 60 ℃ for 30 s and 72 ℃ for 30 s. Expressions of RORC, IL-17, IL-21, IL-6, TGF-β, IL-2 and Foxp3 were evaluated as relative ratios against expression of GAPDH.
Statistical analysis
Following RT-PCR analysis, we evaluated the significant changes of relative mRNA levels for RORC, IL-17, IL-21, IL-6, TGF-β, IL-2 and Foxp3 among patients according to clinical stage and tumor size by using one-way analysis of variance (ANOVA) , and nodal status by using Student’s T-test. Data for one-way ANOVA and Student’s T-test were represented as mean ± SEM. Correlative data for RORC vs. IL-17F, RORC vs. IL-21, RORC vs. Foxp3, RORC vs. IL-6, RORC vs. TGF-β and RORC vs. IL-2 were evaluated by linear least-squares regression analysis. Values of p<0.05 were considered statistically significant. Statistical analysis was performed using a computerized statistics software (JMP14) .
Table 1 Patients and tumor characteristics (n=50)

RORC gene expression in tumor increased with progression of clinical stage in patients with breast cancer
First, we examined expression of the RORC gene, as the master gene of Th17 cells, in breast cancer tissues. RORC mRNA levels in the main tumor of patients with breast cancer were significantly elevated with progression to stage IIIA (Fig. 1A) . RORC expression in the main tumor also increased significantly with tumor growth from pT1 to pT3 (pT1, 20 stage I and 4 out of 10 stage IIA; pT2, 6 out of 10 stage IIA and 10 stage IIB; and pT3, identical to 10 stage IIIA) (Fig. 1B) . However, RORC expression in the main tumor showed no significant difference between nodal status pN0 and pN1 (pN0, 20 stage I and 6 out of 10 stage IIA; and pN1, 4 out of 10 stage IIA, 10 stage IIB and 10 stage IIIA) (Fig. 1C) . These data are concordant with the notion that the increase in Th17 cells correlated with clinical stage, particularly tumor size, in in the main tumor of patients with breast cancer.
RORC expression in SLNs increased with lymph node metastasis
We examined also RORC expression in SLNs, as the first site of lymph node invasion. Although RORC expression generally increased with clinical stage, the pattern of increase for RORC in SLN did not precisely resemble that in the main tumor: RORC expression in patients with stage IIB and stage IIIA significantly increased compared to that in patients with stage I and/ or stage IIA (Fig. 2A) . Furthermore, RORC expression in SLNs showed no significant difference among tumor size statuses (Fig. 2B) , but did show the difference between nodal statuses pN0 and pN1 (Fig. 2C) . These data suggested that the increase in Th17 cells, as judged by RORC expression, in SLNs occurred with lymph node metastasis.
IL- 17 expression in main tumor increased with progression of clinical stage
Next, we examined mRNA levels of IL-17, which is produced mainly by a small group of T cells such as Th17 cells, and which is known as a mediator of tumor progression. IL-17 expression in the main tumor increased with the progression of clinical stage (Fig 3A) , resembling RORC expression (Fig. 1A) . The difference in IL-17 expression was significant among tumor size statuses (Fig. 3B) and nodal statuses (Fig. 3C) , again resembling RORC expression. In analysis by linear least-squares regression, a strong correlation between expressions of RORC and IL-17 in the main tumor was detected (Fig. 3D) . mRNA levels of IL-21, another cytokine up-regulated in Th17 cells, were also elevated in the main tumor significantly with tumor growth from pT1 to pT2 and pT3 (Fig. 3E) . These data are concordant with the notion for the growth and maturation of Th17 cells in the tumor microenvironment of breast cancer.
Tumor-specific cytokine balance sustains the increase in Th17 cells in the tumor microenvironment of breast cancer patients
We evaluated TGF-β and IL-6, which are required for the development of Th17 cells, in the main tumor of breast cancer patients. In the main tumor, mRNA levels of TGF-β were elevated with tumor growth (Fig. 4A) . Furthermore, expression of TGF-β correlated strongly with that of RORC in the main tumor of breast cancer patients (Fig. 4B) . Moreover, mRNA levels of IL-6 were elevated with tumor growth (Fig. 4C) , and strongly correlated with that of RORC in the main tumor of breast cancer patients (Fig. 4D) . These data suggested that TGF-β and IL-6 were correlated with Th17 cells in the tumor microenvironment of progressing tumor in patients with breast cancer.
Next, we checked factors suppressing the development of Th17 cells. We evaluated expression of the Foxp3 gene, as the master gene of Treg with actions opposing Th17 cells in normal immunity. mRNA levels of Foxp3 in the main tumor were also rather elevated with tumor growth from pT1 to pT2 and pT3 (Fig. 5A) , and did not correlate with expression of RORC (Fig. 5B) . Finally, we evaluated expression of IL-2, which is known as a suppressive mediator of Th17 cells. In the main tumor, mRNA levels of IL-2 did not significantly change with tumor growth (Fig. 5C) , and did not correlate with expression of RORC (Fig. 5D) . These results suggested that Treg and IL-2 did not efficiently suppress Th17 cells in the tumor microenvironment of progressing breast cancer.
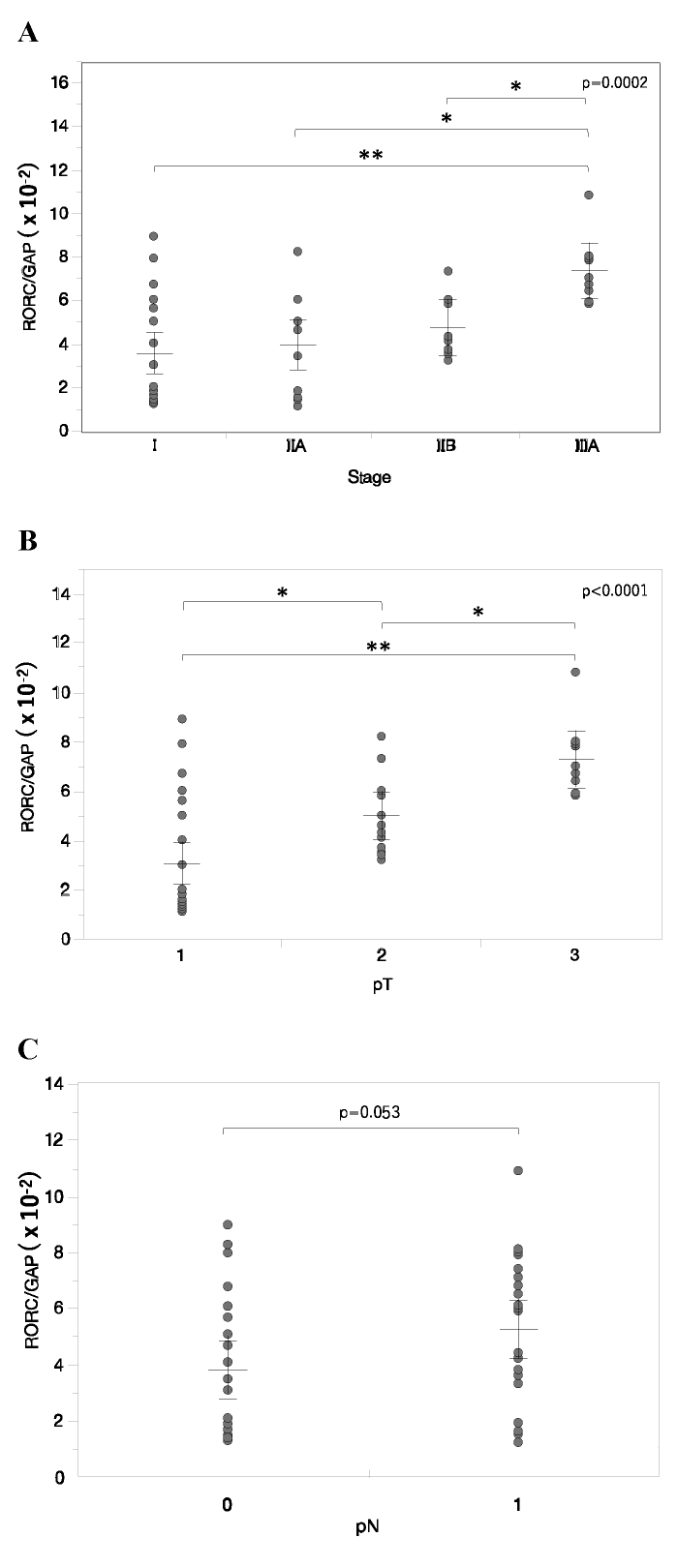
Fig. 1 RORC gene expression in the main tumor of breast cancer. A-C, RORC mRNA levels in the main tumor of varying clinical stages (A) , tumor sizes (B) and nodal statuses (C) . Relative mRNA levels are represented as mean ± SEM. P-value was calculated by one-way ANOVA (A, B) or Student’s T-test (C) . *p<0.05, **p<0.01 by post-hoc test (Tukey-Kramer method) .
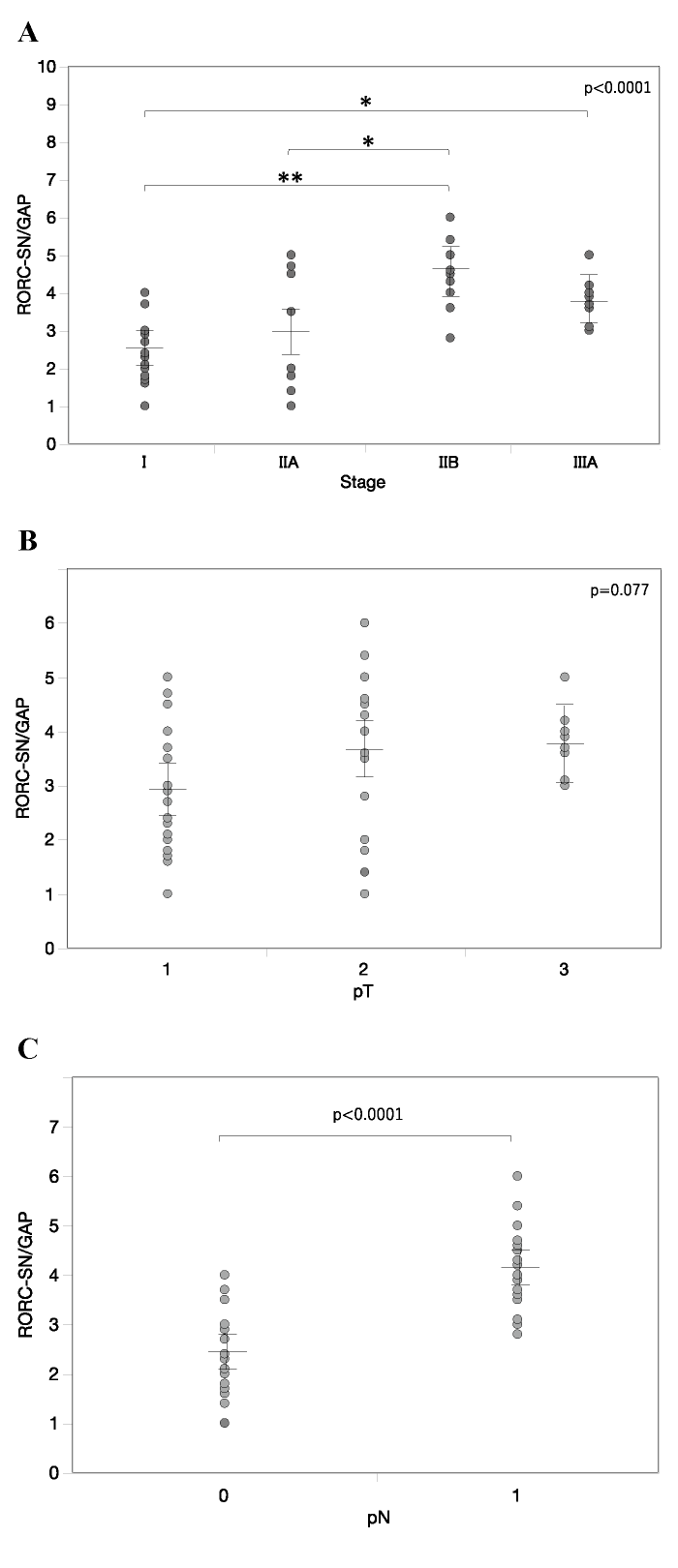
Fig. 2 RORC gene expression in SLNs with the occurrence of lymph node metastasis. A-C, RORC mRNA levels in SLNs of varying clinical stages (A) , tumor sizes (B) and nodal statuses (C) . Relative mRNA levels are represented as mean ± SEM. P-value was calculated by one-way ANOVA (A, B) or Student’s T-test (C) . *p<0.05, **p <0.01 by post-hoc test (Tukey-Kramer method) .
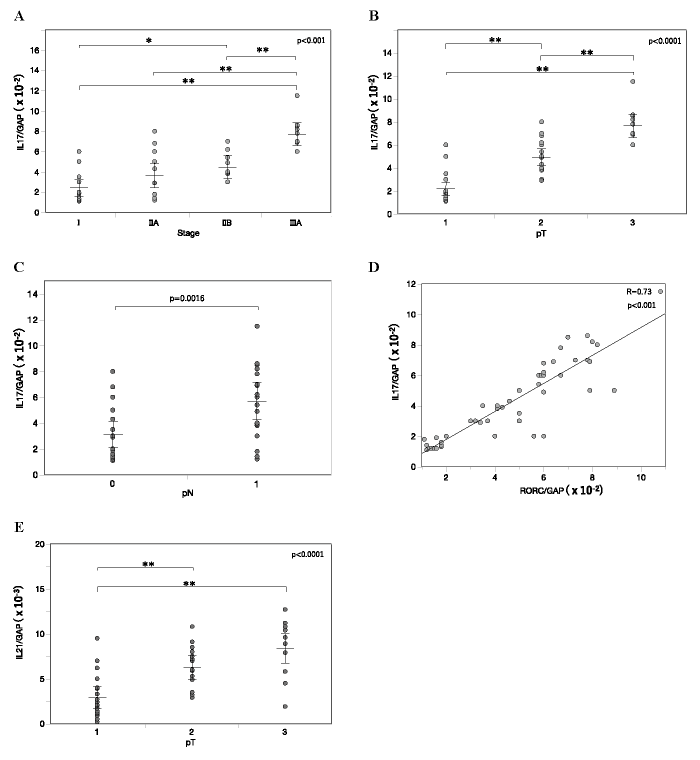
Fig. 3 IL-17 and IL-21 gene expression in the main tumor. A-C, IL-17 mRNA levels in the main tumor of varying clinical stages (A) , tumor sizes (B) and nodal statuses (C) . D, Linear least-squares regression analysis between mRNA levels of RORC (Fig. 1) and IL-17 in the main tumor. E, IL-21 mRNA levels in the main tumor of varying tumor sizes. In (A-C, E) , Relative mRNA levels are represented as mean ± SEM. P-value was calculated by one-way ANOVA (A, B, E) or Student’s T-test (C) . *p<0.05, **p<0.01 by post-hoc test (Tukey-Kramer method) .
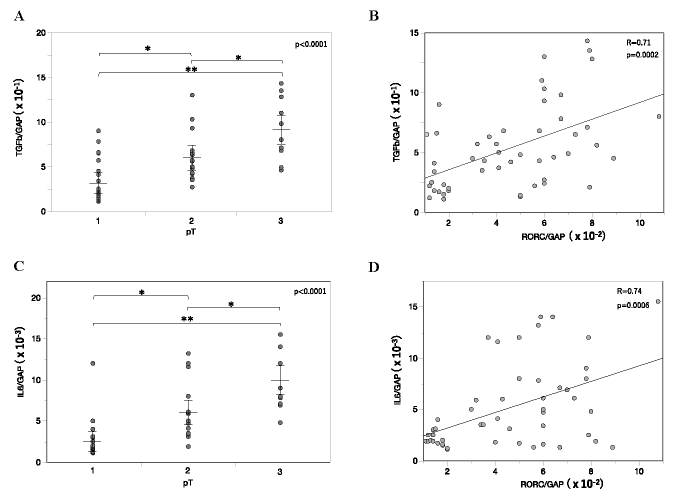
Fig. 4 TGF-β (A, B) and IL-6 (C, D) gene expression in the main tumor. A, C, TGF-β (A) and IL-6 (C) mRNA levels in the main tumor of varying tumor sizes. Relative mRNA levels are represented as mean ± SEM. P-value was calculated by one-way ANOVA. *p<0.05, **p<0.01 by post-hoc test (Tukey-Kramer method) . B, D, Linear least-squares regression analysis between mRNA levels of RORC (Fig. 1) and TGF-β (A) or IL-6 (C) .
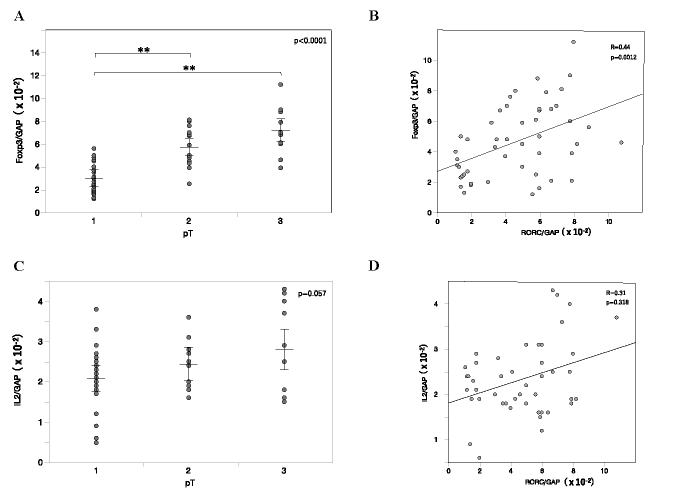
Fig. 5 Foxp3 (A, B) and IL-2 (C, D) gene expression in the main tumor. A, C, Foxp3 (A) and IL-2 (C) mRNA levels in the main tumor of varying tumor sizes. Relative mRNA levels are represented as mean ± SEM. P-value was calculated by one-way ANOVA. **p<0.01 by post-hoc test (Tukey-Kramer method) . B, D, Linear least-squares regression analysis between mRNA levels of RORC (Fig. 1) and Foxp3 (B) or IL-2 (D) .
Cancer and inflammation have long been considered two sides of the same coin[4,5]. Due to advances in the understanding of the tumor microenvironment, cell groups and surrounding cytokines associated with tumor-specific inflammation have been brought back into the spotlight for the purposes of screening for new therapeutic targets[1-3]. The function of the longterm maintenance of tumor-specific inflammation during tumor progression, such as the mechanisms and platform for continuous production of tumor-specific inflammatory cytokines, remains unclear. In particular, IL-17 has been known as an inflammatory cytokine and was thought to be produced by Th1 cells[21,36]. A previous study showed that Th2 cells are dominant over Th1 cells in the tumor environment [37,38]. Although TGF-β and IL-6 were produced by tumor [39-42], increases in the Th1 cytokine IL-17 were difficult to explain the tumor microenvironment [29].
With the clarification of Th17 cells as a subset of CD4 (+) T cells producing IL-17[9-11], the relationship between this cell subset and tumor-specific inflammation in the tumor microenvironment gained attention. Kryczek et al. reported a clear prevalence of Th17 cells in mouse and human cancers[25]. Zhang et al. described an increase in Th17 cells with tumor progression of human gastric cancer [28]. The present data suggested that Th17 cells, at least in terms of marker mRNA levels, increased with tumor progression in the main tumor and SLNs. In the main tumor, expression of the RORC gene, as the master gene of Th17, increased with tumor growth. In SLNs, expression of the RORC gene was seen with the development of lymph node metastases. Kryczek et al. reported that the priming site of Th17 cells is not drainage lymph nodes [25]. Similarly, our data suggested that priming of Th17 cells was generated within or surrounding the tumor, not in SLNs, because RORC gene expression in SLNs from patients with pN0 was not increased with progression of the main tumor (data not shown) . Furthermore, the present study provided a circumstantial evaluation of the balance of tumor-specific inflammatory cytokines with tumor progression in patients with breast cancer. Along with the progression of breast cancer, IL-17 gene expression increased with RORC gene expression, being likely that IL-17 is produced by Th17 cells in the tumor microenvironment of breast cancer. Furthermore, TGF-β and IL-6 gene expressions correlated closely with RORC gene expression. Kryczek et al. reported that Th17 cells were coexistent with Treg cells, which are known as suppressors of Th17 cell development, and the balance of Th17 and Treg cells was regulated by IL-2 in the tumor microenvironment [25]. In our data, IL-2 gene expression did not increase, while TGF-β and IL-6 gene expressions increased with tumor progression in breast cancer patients. Although Treg cells may be increased by tumor-produced TGF-β[14], Th17 cells may be predominantly produced by the tumor-specific cytokine milieu with the progression of breast cancer. These data suggest that the development of Th17 cells is accelerated in the tumor microenvironment of breast cancer, IL-17 produced by Th17 cells leads to the increase of TGF-β and IL-6, and finally a malignant cycle maintaining tumor-specific inflammation may be built up.
Muranski et al. reported that tumor-specific Th17- polarized cells could eradicate large established melanomas in a mouse model [26]. Furthermore, Sfanos et al. reported that increased Th17 cells in the prostate showed an inverse association with Gleason score, an indicator of the malignancy of prostate cancer [27]. Whether Th17 cells play a key role in antitumor immunity or, inversely, a central role in tumor progression by maintaining tumor-specific inflammation, remains unclear. Our data implicitly support the latter possibility, as Th17 cells increased with clinical stage of breast cancer. Future investigations will hopefully clarify the relationship between increased Th17 cells in tumor and prognosis for breast cancer.
In summary, biomarker expression for Th17 cells increased with tumor progression, likely affecting the tumor-specific cytokine balance in the tumor microenvironment of breast cancer. We presumed that in the malignant cycle of tumor progression and inflammation maintenance, Th17 cells may play a key role in tumor progression by maintaining tumor-specific inflammation. Th17 cells and the surrounding cytokines associated with tumor-specific inflammation may offer a new therapeutic target for breast cancer.
MA and MS conceived and designed the study. MA, MS and TN were involved in data collection. MA and MS analyzed and interpreted the data. MA, MS and TN were involved in the development and approval of the manuscript. MO oversaw the whole process of writing the manuscript.
None to declare.
MO is a member of the Editorial board of the Chiba Medical Journal. MA, MS, and TN declare that they have no conflicts of interest, either financial or non-financial, with regard to the content of this article.
Consent for sampling in this study was based on the study of clinical biobank at Chiba University with the approval of Research Ethics Committee of Graduate School of Medicine, Chiba University (Bioethics review number #942) . Written informed consent was obtained from all individuals for the present study protocols.
All data are available in the manuscript, and Excel files may be obtained from the corresponding author upon request.
Address correspondence to Dr. Masahiro Sakakibara.
Departments of Breast Surgery, Toho University Sakura Medical Center,
564-1, Shimoshizu, Sakura, Chiba 285-8741, Japan.
Phone: +81-43462-8811.
E-mail:masahiro.sakakibara@med.toho-u.ac.jp
