




Chiba Medical J. 99E:1-7, 2023
doi:10.20776/S03035476-99E-1-P1
〔 Chiba Medical Society Award Review 〕
Hirokazu Usui
Department of Reproductive Medicine, Chiba University Graduate School of Medicine, Chiba 260-8670.
(Received September 30, 2022, Accepted November 9, 2022, Published February 10, 2023.)
Hydatidiform moles are abnormal villous diseases characterized by excessive trophoblast proliferation. They are classified into two categories: complete hydatidiform moles (CHM) and partial hydatidiform moles (PHM), whose cytogenetic constitution is exceptional. Gestational trophoblastic neoplasia (GTN) can develop from both types of hydatidiform moles. The incidence of GTN is different between CHM and PHM. However, it is occasionally difficult to diagnose hydatidiform moles. Thus, we have continued to diagnose them using molecular cytogenetic procedures, such as short tandem repeat (STR) polymorphisms, to determine their critical diagnosis. Based on STR analysis of CHM and PHM, the incidence of GTN was 15% and 2%, respectively. Furthermore, single nucleotide polymorphism array analysis has been introduced into the molecular karyotyping of whole genomes. Single nucleotide polymorphism array analysis clarified that heterozygous androgenetic CHM is of dispermic origin, but not a diploid sperm. Additionally, we showed that one-third of heterozygous androgenetic CHM had trisomic chromosomes. The origins of the additional chromosomes were not only paternal but also maternal, although they should be androgenetic. Molecular karyotyping revealed a spermatogonium-derived complete hydatidiform mole. A distinct diagnosis of hydatidiform moles is important for management of hydatidiform moles. Molecular biological procedures could elucidate the mechanisms underlying the development of hydatidiform moles.
B allele frequency, hydatidiform moles, molecular karyotyping, short tandem repeat, single nucleotide polymorphism array
Hydatidiform mole (HM) is a disease characterized by excessive proliferation of trophoblasts, which can be diagnosed by histopathological findings (Fig. 1) [1,2]. Hydatidiform moles are classified into two categories: complete hydatidiform moles (CHM) and partial hydatidiform moles (PHM) . A unique cytogenetic constitution of CHM was reported in 1977 by Kajii and Ohama[3]. The CHMs were androgenetic and lacked maternal chromosomes. The cytogenetic constitution of the PHMs was shown to be triploid (Fig. 2) [4,5]. Complete hydatidiform moles and PHM are occasionally difficult to diagnose pathologically[6,7]. These characteristics have been utilized for auxiliary diagnostic techniques, such as karyotyping, enzyme polymorphism assays, flow cytometry, fluorescence in situ hybridization, immunohistochemistry of p57KIP2, and short tandem repeat (STR) polymorphism analysis [8-12].
The treatment and management of HMs are fundamental because they have a potential for malignant transformation to gestational trophoblastic neoplasia (GTN) [1,13]. We have continued to specialize in the treatment and research of trophoblastic disease[6,14-19]. Molar pregnancies and trophoblastic tumors are rare with approximately 2,000 and 200 respective cases per year in Japan[20]. We conducted basic clinical research based on the genetic diagnosis of HM. More than 500 cases of HMs have been genetically examined [14,15,21].
In this review, we introduce the results of our research using DNA polymorphism analysis of HMs, particularly new findings on the pathogenesis of HMs, as revealed by recent single nucleotide polymorphism (SNP) analysis.
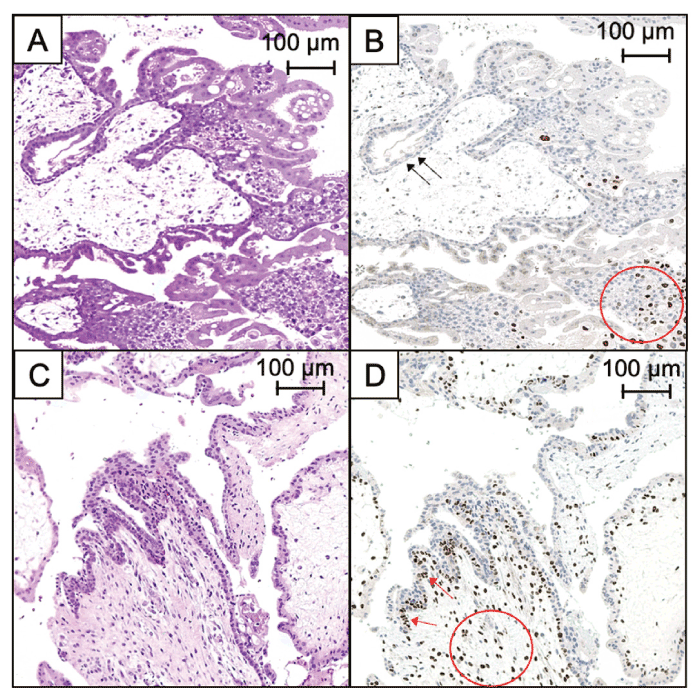
Fig. 1 Pathological features of hydatidiform moles. Complete hydatidiform moles (CHM). (A) Hematoxylineosin staining (× 100, scale bar: 100 μm); (B) immunohistochemistry of p57KIP2. Extravillous trophoblasts were stained (red circles) (× 100, scale bar: 100 μm). Cytotrophoblasts and stromal cells of CHM were not stained with anti-p57KIP2 antibody (black arrows). Partial hydatidiform moles (PHM). (C) Hematoxylin-eosin staining (× 100, scale bar: 100 μm). (D) Immunohistochemistry of p57KIP2 (× 100, scale bar: 100 μm). Cytotrophoblasts (red arrows) and stromal cells (red circles) of PHM were stained with anti-p57KIP2 antibody.
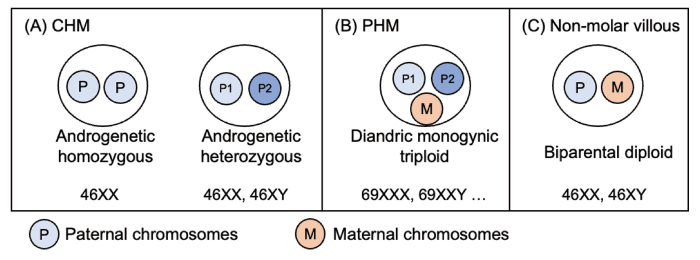
Fig. 2 Cytogenetic characteristics of hydatidiform moles. (A) Complete hydatidiform mole (CHM) is an androgenetic diploid, classified into two types: homozygous CHM, which results from the duplication of a haploid sperm; and heterozygous CHM, which results from the fertilization of two mature sperms. (B) Partial hydatidiform mole (PHM) is diandric monogyny triploid. (C) Non-molar villous trophoblasts are biparental diploids. (Figure 2 was adapted from reference No. 8 by Usui, 2022.) CHM, complete hydatidiform mole; PHM, partial hydatidiform mole
Recently, STR polymorphism analysis has become the gold standard auxiliary diagnostic method for HMs, which can be used for identification in forensic medicine[2,8]. Short tandem repeat polymorphism analysis can be used to distinguish between CHM, PHM, and non-molar pregnancies. Multiplex-STR kits are commercially available and they include 16 or more polymorphic loci in a single reaction[22]. Additionally, single electrophoresis can yield multi-fragment charts in multi-fluorescence colors.
Typical examples of STR polymorphism analysis are shown in Figure 3[14,23]. Non-molar pregnancies are diploid in origin from both parents, and the villous locus inherits one allele each from the mother and father. Complete hydatidiform moles are androgenetic, and the villous locus does not contain mother-derived alleles. Furthermore, it is possible to distinguish between homozygous and heterozygous CHMs. The villous loci of PHMs consist of two alleles of paternal origin and one allele of maternal origin. When the villous alleles were two, the peak ratio was 2:1, resulting in a consistent result for two paternal and one maternal allele. From fresh surgical specimens, it took 4 h for genome extraction, 2 h for STR polymerase chain reaction (PCR), and 3 h for electrophoresis. The following day, the results were obtained.
Short tandem repeat analysis is a reliable technique; however, facilities cannot perform STR analysis routinely. Immunohistochemical detection of p57KIP2 can be used to detect androgenetic CHM[11,24,25]. The cytotrophoblasts and stroma cells of androgenetic CHM were not stained using p57KIP2 antibody, while those of PHM and biparental abortion were stained using p57KIP2 antibody (Fig. 1).
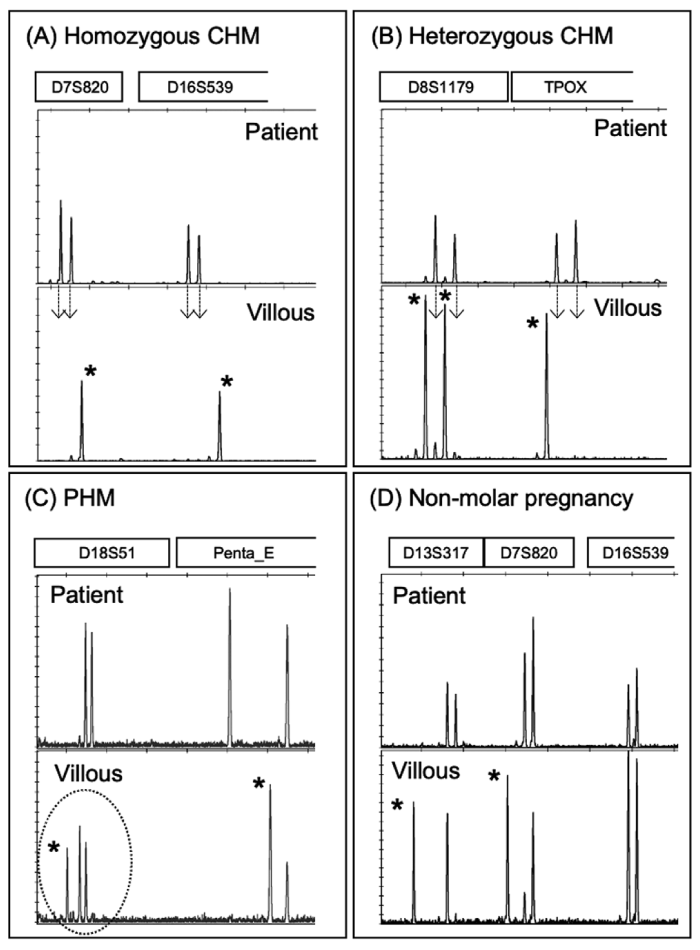
Fig. 3 Short tandem repeat polymorphism analysis of hydatidiform moles. Loci of the patient and villous tissues of the androgenetic complete hydatidiform moles (CHMs) are shown. (A) Androgenetic homozygous CHM, (B) androgenetic heterozygous CHM. Dotty arrows indicate the two maternal alleles that were not entirely inherited in the villi. (C) Some loci of the partial hydatidiform mole had three allelic loci, indicating that they would have three chromosomes. (D) Non-molar villous tissue showing biparental diploidy. Asteristics indicate estimated alleles of paternal origin. CHM, complete hydatidiform mole; PHM, partial hydatidiform mole
As described above, CHM and PHM differ in clinical and biological characteristics. In clinical studies related to HMs, a lack of complete distinction between CHM and PHM leaves ambiguity in the results. Therefore, we conducted a clinical study of trophoblastic diseases based on the diagnosis using STR polymorphism analysis.
First, we conducted a prospective cohort observational analysis to investigate the incidence and risk factors of GTN in cytogenetically diagnosed HMs from 2007 to 2016. The incidence of GTN from androgenetic CHM, triploid PHM, and non-molar biparental pregnancy were 15.1% (35/232), 1%, and 0% (0/88), respectively. One patient with cytogenetic PHM developed GTN, although the risk of developing GTN was lower for PHMs than for CHMs [14].
Patients with low-risk GTN need to receive chemotherapy. Methotrexate (MTX) is a standard drug used for low-risk GTN. Low-risk GTN is sensitive to treatment with MTX. However, almost half of the patients did not achieve remission after MTX treatment. The primary target of MTX is folate metabolism. Methylenetetrahydrofolate reductase (MTHFR) is an essential rate-limiting enzyme involved in folate metabolism. Methylenetetrahydrofolate reductase polymorphism C677T is a functional SNP with amino acid changes and decreased enzyme activity. Thus, we performed an association study of two representative MTHFR polymorphisms, C677T and A1298C, in patients and CHM villous tissues. The MTHFR 677T allele in molar tissue was related to the need for drug change (odds ratio[OR], 3.13; 95% confidence interval[CI], 1.31–7.49; P=0.006), although the MTHFR 677T alleles obtained from patient’s blood DNA were not related to the outcome of MTX treatment (OR, 1.31; 95% CI, 0.61–2.91; P=0.49) . These data demonstrate that the MTHFR 677TT genotype in molar tissue is associated with failure of MTX treatment in low-risk patients with GTN[26].
The conceptus is genetically different from its mother. Complete hydatidiform moles are allografts for patients with an androgenetic origin. Therefore, we hypothesized that ABO-compatible cases between molar tissues and patients could easily develop into GTN because CHM can escape immunological rejection. We evaluated the effect of ABO blood-group compatibility on the prognosis of androgenetic CHMs. We determined the molar ABO blood types by SNP genotyping using a high-resolution melting assay. According to ABO blood group incompatibility between patients and molar tissues, GTN occurred in 19.1% (18/94) of compatible cases and 11.4% (4/35) of incompatible cases; the incidence did not differ statistically (P=0.223) . The compatibility of ABO blood group between patients and molar tissues had no relationship with the appearance of GTN[27]. GTN occurred in 10.8% (4/37), 14.8% (8/54), 22.2% (6/27), and 36.4% (4/11) of patients with types O, A, B, and AB blood groups, respectively. Patients with blood type AB are more likely to develop GTN[27].
We introduced SNP array analysis to evaluate HMs[21,23,28,29]. The SNP array has a substantial advantage in that it can cover whole genome information. A large number of loci can overcome the binary characteristics of SNPs and determine molecular karyotyping. The signal intensity reflects the copy number, enabling viewing chromosomal or region loss or gain. Allelic differences may indicate loss of heterozygosity. The B allele frequency (BAF) was normalized to reflect the allelic intensity ratio of the A and B alleles. A BAF of 0 or 1 indicates that one of the two alleles (AA or BB) is absent, whereas a BAF of 0.5 means that both alleles are equally distributed (AB) . BAF plots on ideograms can be utilized for molecular karyotyping, which is appropriate for diagnosing HMs. (Fig. 4) [23]. Androgenetic CHMs showed the homozygosity in all regions for homozygous CHM (Fig. 4A), and in part (mixed homozygous and heterozygous) for heterozygous CHM (Fig. 4B) . Partial hydatidiform moles should have four regions (0, 0.33, 0.66, 1.0 for AAA, AAB, ABB, and BBB locus, respectively) . Otherwise, non-molar villi are all heterozygous, showing plots of BAF 0, 0.5, and 1.0, according to AA, AB, and BB allele over the whole chromosomes.
Heterozygous CHM is sometimes referred to as a dispermic mole because its chromosome composition does not contradict its dispermic origin. However, both mechanisms fertilizing two sperms or a diploid sperm could be postulated, and there is no direct evidence for them. We analyzed androgenetic heterozygous CHMs using BAF plots of SNP arrays. In cases of diploid sperm fertilization, the centromere status would show all heterozygous (meiosis I error) or all homozygous (meiosis II error), depending on their pathogenesis [23]. However, the centromeric zygosities showed both homozygous and heterozygous statuses in all the analyzed cases, indicating two sperm fertilizations [23].
The BAF plot can detect aneuploid chromosomes more easily than copy number abnormalities, such as gain or loss of genes. Triploid chromosomes would show the four regions at AAA, AAB, ABB, and BBB in the BAF plot. One-third of androgenetic heterozygous CHM (dispermic) (9/31) had trisomic chromosomes. One of these has double trisomic chromosomes [21].
We developed a selected BAF plot, which could visualize the origin of chromosomes, as paternal or maternal. Surprisingly, the additional chromosomes, six of the nine heterozygous aneuploid CHMs, were of maternal origin, although the androgenetic CHMs were of paternal origin (Fig. 5) . We estimated that there might be three pronuclear zygotes including one oocyte and two sperms nucleus, as the intermediates of androgenetic heterozygous CHMs. Furthermore, the incidence of GTN in heterozygous androgenetic CHM, with or without maternal chromosomes, was 0% (0/6) and 28% (7/25), respectively. The relationship between the development mechanism and malignant potential of CHMs should be studied in the future[21].
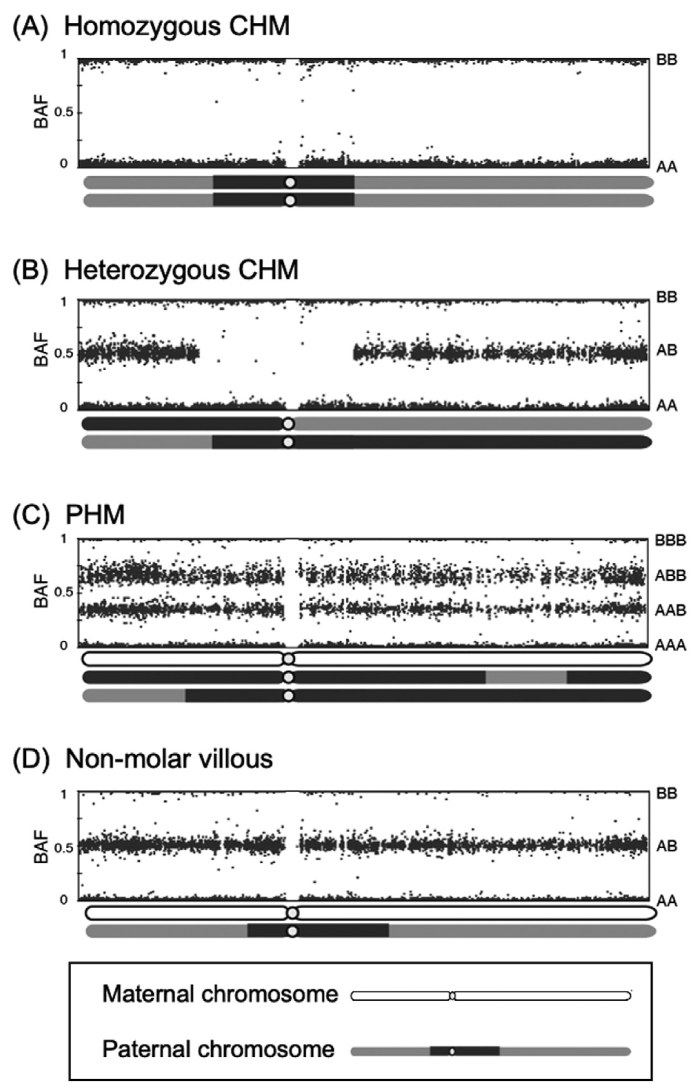
Fig. 4 B allele frequency plots from SNP array data for hydatidiform moles. Chromosome constitutions and corresponding B allele frequencies are shown for: (A) an androgenetic homozygous complete hydatidiform mole; (B) an androgenetic heterozygous complete hydatidiform mole; (C) a diandric monogynic triploid partial hydatidiform mole; and (D) biparental diploid non-molar villi. (Figure 4 was adapted from reference No. 23 by Usui et al., 2019.). CHM, complete hydatidiform mole; PHM, partial hydatidiform mole; BAF, B allele frequency
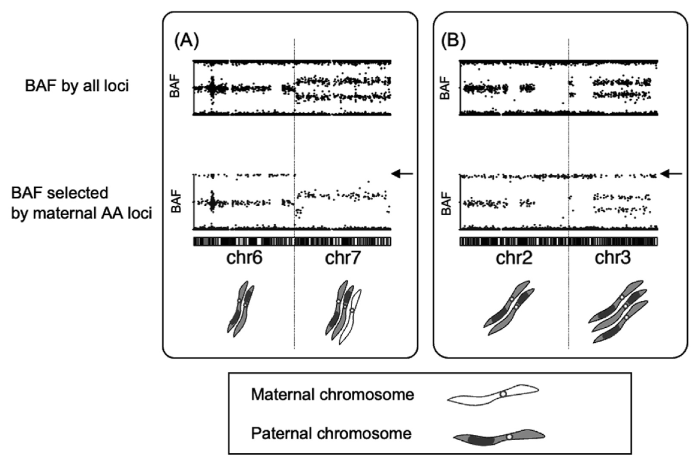
Fig. 5 Determination of parental origin of additional chromosomes of androgenetic heterozygous complete hydatidiform moles. (A) Upper chart: chromosomes 6 and 7 showing diploidy and trisomy, respectively. Lower chart: BB regions are present on chromosome 6, but not on chromosome 7, indicating that chromosome 6 is androgenetic diploid and chromosome 7 is diandric monogyny triploid. (B) Upper chart: chromosomes 2 and 3 showing diploidy and trisomy, respectively. Lower chart: BB regions are present on both chromosomes 2 and 3, which indicates that chromosome 2 was androgenetic diploid, and that three chromosomes of chromosome 3 were all of paternal origin. (Figure 5 was adapted from reference No. 21 by Usui et al., 2020.)
Recently, we analyzed androgenetic CHM in a woman who underwent intrauterine implantation of a blastocyst obtained by in vitro injection of a presumed round spermatid into her oocyte. The origin of androgenetic CHM was the spermatogonium rather than the round spermatid[29]. This CHM did not contain maternally derived nuclear DNA, but contained mitochondrial DNA. Furthermore, the CHM genome is genetically identical to that of paternal blood DNA.
This study indicated that spermatogonium, which should have identical genomes with somatic cells, could potentially develop into androgenetic CHM.
We reviewed the results of our study, mainly focused on androgenetic CHMs. We performed clinical and basic research on trophoblastic disease using STR-PCR and SNP array analysis on patients and villous genomes. The development mechanism of androgenetic heterozygous CHMs is of dispermic origin but not diploid sperm. Three pronuclear zygotes might be the first step in heterozygous androgenetic CHM. The selected BAF plotting procedure was successfully utilized for molecular karyotyping. Further studies are required to elucidate the mechanisms underlying the development of HMs.
This work was partly supported by the Japan Society for the Promotion of Science KAKENHI (grant numbers JP 21K19551 and 22H03220) .
The author declares no conflict of interest associated with this manuscript.
Not applicable.
Not applicable.
The author expresses gratitude to Prof. Makio Shozu for critically reading the manuscript.
Address correspondence to Dr. Hirokazu Usui.
Department of Reproductive Medicine, Chiba University
Graduate School of Medicine, 1-8-1, Inohana, Chuou-ku,
Chiba 260-8670, Japan.
Phone: +81-43-226-2122.
Fax: +81-43-226-2121.
E-mail: hirokazu-usui@faculty.chiba-u.jp
