| School
of Medicine, Chiba University Chiba University Hospital |
|
|
|
|
|
| Chiba Medical Journal | |
| Journal Index | |
| 〔Original Paper〕 | ||||
| Gaze recognition in high-functioning autistic patients: evidence from functional MRI |
||||
|
|
||||
| Hiroko Takebayashi1), Hideo Matsumoto2), Masahiro Ogai1), Yoshimoto Sekine1) Katsuaki Suzuki1), Hideo Matsuzaki1), John Suckling3), Fukujirou Ozawa4) Rinmei Fukuda5), Ichiro Uchiyama6), Kazuhiko Nakamura1), Yoshio Minabe1) Haruo Isoda7), Norio Mori1) and Nori Takei1, 8) | ||||
| (Received November 29, 2005, Accepted November 30, 2005) | ||||
|
|
||||
SUMMARY
|
||||
Key Words
|
||||
| Ⅰ.Introduction Autistic disorder (AD) is characterized by a dysfunction of communication and impaired development with respect to the acquisition of social skills. Some authors have suggested that AD is a condition involving an impaired “Theory of Mind” (TOM) or mentalizing (Baron-Cohen et al[1]; Happe and Frith[2]; Castelli et al[3]). TOM refers to the ability to perceive the mental states of other persons, such as the beliefs and intentions of others; that is, with this function, one can understand and predict the behavior of other people by reasoning about their mental states (Premack and Woodruff[4]). An abundance of information derived from the expressions on the human face, and face-to-face communication is vital in interpersonal interactions. Using neuroimaging technology, some groups have focused on human faces as stimuli to investigate TOM function in AD patients. Three main approaches have been applied, i.e., face recognition, facial expression, and eye gaze experiments. For example, as regards the face recognition, Schultz et al[5]]have revealed using functional magnetic resonance imaging (fMRI) that AD patients exhibit greater activation in the inferior temporal gyri on a face discrimination task than do control subjects. Since the activated areas were generally involved in the completion of non-face object perception tasks, their results suggested that AD patients may process human faces merely as objects. A study reported by Critchley et al[6]demonstrated widespread functional abnormalities in the limbic region, the cerebellum, and the extrastriate visual cortices during the explicit and implicit processing of emotional facial expressions in individuals with AD. In our previous fMRI study, we observed differential brain function between patients with AD and normal controls during tasks involving the recognition of facially expressed emotions (Ogai et al[7]). Baron-Cohen et al[8], who studied brain function in autistic patients and normal subjects using a task involving the recognition of a mental state expressed in another person’s eyes, reported less activation in the amygdala and more activation in the superior temporal gyrus in the AD patients than in the normal subjects. Baron-Cohen[9]has also suggested that the interpretation of another person’s gaze plays an important role in a normally functioning TOM. In accord with this suggestion, some functional imaging studies of normal volunteers have shown that tasks involving the TOM, as well as tasks involving eye gaze processing, evoked activation in similar brain regions, such as the posterior superior temporal sulcus (Hoffman and Haxby[10]), the medial frontal areas (Calder et al[11]), and the amygdala (Kawashima et al[12]). Therefore, the presentation of gaze discriminating tasks as a social stimulus can be considered to be sufficient to activate the mechanisms by which individuals process and interpret the mental states of others. We tested the hypothesis that on an eye-gaze paradigm in the framework of fMRI, AD patients with a relatively high IQ (i.e., individuals with high-functioning AD) would show abnormal activation in brain regions implicated in the functioning of TOM. Ⅱ.Materials and Methods 1.Subjects We studied 5 right-handed patients satisfying the DSM-IV (American Psychiatric Association[13])criteria for AD and 9 right-handed normal subjects who were matched with the patients in terms of age, IQ (both groups had an IQ of above 85 on the full Wechsler Adult Intelligence Scale Revision, WAIS-R), educational level, and socioeconomic status (Table 1). Diagnosis of the patient group was made in an operationalized manner identical to that of the Autism Diagnostic Interview-Revised (ADI-R; Lord et al[14]). The patient group consisted of individuals with high-functioning AD. No comorbidity of psychiatric problems other than AD was identified in any of the subjects. None of the patients were medicated at the time of the study. After providing the subjects with a complete description of the study, written informed consent was obtained from all participants before entry into the study, in compliance with the procedures of the Ethics Committee of the Hamamatsu University School of Medicine. | ||||
|
||||
|
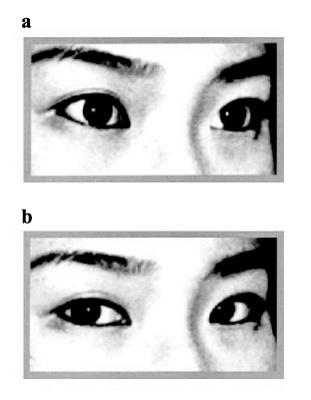
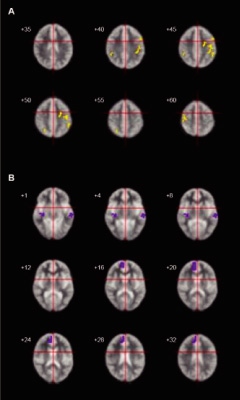
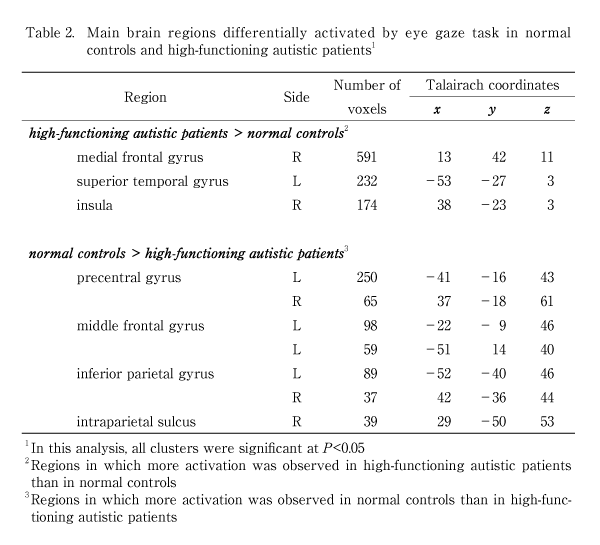
2.Experimental paradigms and procedure As the stimulus during the scanning, we implemented a gaze recognition task. During the task, pictures of other people’s eyes, showing both direct and averted gazes, were presented (Fig. 1). These eye gaze stimuli were counterbalanced for age and gender. The same gaze orientations were used for all stimuli. The person’s head tilted 15° leftward from the front for all the conditions. In the direct condition, the person’s gaze was directed toward the camera. In the averted condition, the person’s gaze was directed 15° leftwards from the front. The subjects were instructed to determine the direction of the gaze. Stimuli were presented through goggles worn by the subjects. Before the MRI scan, the subjects were presented with an example that was not used during the actual scanning time; this example appeared on the computer monitor, and was used to familiarize the subjects with the experimental procedure and the experimental conditions, in order to minimize anxiety and enhance task performance. After this familiarization, the subjects lay supine on the scanning table and were fitted with goggles. We used an alternating on/off box-car design of 30 sec duration for each on or off condition, and there was a total of 5 cycles of “on/off” alternation. Each stimulus was presented for 3.5 sec with an interval of 0.25 sec. In the “on” condition, 8 different faces with a direct gaze were presented to the subjects. In the “off” condition, the same 8 faces were shown with an averted gaze. During the scanning procedure, subjects were asked to concentrate on their judgment of the direction of the gaze. After the scanning session, the subjects immediately performed a test, on the perception of the gaze presented in the experiments. The brain activation reported in this article refers to periodic BOLD (blood oxygenation level-dependent; Ogawa et al[15]) signal changes at the fundamental experimental frequency. 3.Image acquisition One hundred T2-weighted gradient-echo (TR=3000, TE=40 ms) echoplanar MR images depicting BOLD contrast were acquired using a 1.5 Tesla scanner GE Signa system (General Electric, Milwaukee, Wisconsin, USA) for a 5-min period at 14 near-axial non-contiguous 7-mm thick planes (0.7 mm gap) parallel to the intercommissural (AC-PC) line encompassing the whole brain. During the same session, a 43-slice, high-resolution inversion recovery echo planar image of the whole brain was also acquired, again parallel to the AC-PC line, from each subject (TI=180 ms, TE=73 ms, TR=16 s, 1.5 mm plane resolution, 3 mm slice thickness, 8 signal averages) in order to determine the subsequent anatomical registration. 4.Data analysis Data were processed by the Brain Activation and Morphological Mapping software (University of Cambridge, UK: http://www-bmu.psychiatry.cam.ac.uk/software/). Following correction for the effects of subject’s motion during image acquisition (Bullmore et al[16]), linear regression was used to estimate experimentally induced signal changes at each voxel between the direct (“on”) and averted eye gaze (“off”) conditions. Prior to model fitting, this single contrast design matrix was convolved with a pair of Poisson kernels to model locally variable hemodynamics. A test statistic of the standardized power of the response (F*) was computed from the ratio of the sum of the squares due to the model fit and the residual sum of the squares. Statistic maps representing the standardized effect for each individual at all sessions were registered in the standard space of Talairach and Tournoux[17]by an affine transformation to a template image (Brammer et al[18]). Differences in the responses (F*) between AD subjects and normal subjects were calculated at each voxel with a second-level regression of the general linear model. These differences were tested for statistical significance by a two-tailed test against a distribution sampled under a null-hypothesis of no group effect by the repeated permutation of group membership of the subjects and by recalculation of the differences. Due to the large number of voxel locations which were tested (typically~300,000), the estimated number of type I errors at reasonable statistical thresholds (P-values) was unacceptably large. Spatial information (clusters) was included in the test statistics in order to reduce the overall number of tests, in addition to improving sensitivity to large areas of low-contrast differences. In brief, the cluster-level analysis involved applying a preliminary probability threshold (P<0.05) to the observed difference maps and setting all subthreshold voxels to zero, thus creating a set of suprathreshold voxel clusters that were spatially contiguous in 3D. An identical procedure was applied to the maps of the simulated null-hypothesis, with the resulting clusters sampling the null-distribution for the cluster statistic. The family wise-error rate was controlled by rejecting the null-hypothesis with a statistical threshold at which only 1 false positive (type I error) cluster per image was expected by chance (for further details, see Bullmore et al[19]). For behavioral performance, we compared the correct answers in 8 different persons’ eye stimuli of direct and averted gazes, which were used during the actual scanning time, between AD and control groups using t test. Ⅲ.Results 1.Behavioral performance There was no significant difference between the autistic and control groups as regards the proportion of accuracy when the subjects were identifying either of the two types of gaze; rates of 67.5% vs. 68.1% were obtained, respectively, for direct gaze (t=-0.03, P=.98), and 97.5% vs. 100% for averted gaze (t=-1.39, P=.19), respectively. 2.fMRI findings More activation was observed in the controls than in the AD patients in the left middle frontal gyrus, the bilateral precentral gyrus, and the right intraparietal sulcus, as well as in the bilateral inferior parietal lobe (Fig. 2A and Table 2). In contrast, more activation was observed in the patients with AD in the left superior temporal gyrus, the right insula, and the right medial frontal gyrus, as compared to the controls (Fig. 2B). | ||||
|
||||
|
Ⅳ.Discussion Our results demonstrated that the medial frontal gyrus and the superior temporal gyrus were more activated in AD patients on an eye gaze task than in the control subjects. These brain regions are consistent with the areas implicated in the processing of mental states of other people, i.e., those areas involved in TOM (Fletcher et al[20]; Happe et al[21]; Gallagher et al[22]; Stuss et al[23]; Calarge et al[24]). As mentioned above, the process of reasoning about the significance of another person’s gaze has been suggested to involve a mechanism of TOM (Baron-Cohen[9]). Neuroimaging studies have shown in healthy subjects that tasks directly involving the TOM mechanism as well as eye gaze processing tasks induce activity in these two brain regions (Hoffman and Haxby[10]; Calder et al[11]). Therefore, our findings suggest that processing the gaze of others provokes a dysfunction in TOM-related brain regions in high-functioning AD patients. Baron-Cohen et al[8]conducted an fMRI study which focused on the mental state expressed in another person’s eyes, and have found that AD patients show less activation in the left amygdala than normal subjects. However, we failed to detect any difference in activation in the amygdala between the AD and control groups. The failure to demonstrate reduced activation in the amygdala (Baron-Cohen et al[8])in our sample of AD patients could have been due to differences in the tasks employed between the two studies. Our task focused on the judgment of the direction of the other person’s gaze, whereas the task used in the study by Baron-Cohen et al[8]incorporated a determination of the complex attributes of emotions expressed in the eyes of others. Although it has been suggested in a previous fMRI study that emotional responses are evoked during the performance of eye gaze tasks in normal volunteers (Kawashima et al[12]), our study did not specifically ask the subjects to identify the mental state expressed in the other person’s eyes. This difference may account for the lack of a functional impairment in the amygdala in our sample of AD patients. We also found different patterns of activation between AD patients and the control subjects in other brain regions such as the insula, the precentral gyrus, the middle frontal gyrus, and the intraparietal sulcus; these four regions have not been previously indicated to be related to the functioning of TOM, and therefore these findings were not anticipated. Our results revealed that AD patients showed more activation in the insula compared to the control subjects, whereas there was less activation in the precentral gyrus, the middle frontal gyrus, and the intraparietal sulcus in the AD patient group than in the control group. It has been suggested that the insula becomes involved when stimuli directly evoke strong emotional responses, such as disgust (Adolphs et al[25]; Morris et al[26]; Phillips et al[27]). The precentral gyrus has been shown to be associated with visual search processing (Darren et al[28]; Gitelman et al[29]), and patients with AD have been reported to engage more heavily in visual searching on face perception tasks, whereas healthy controls tend to utilize more holistic processing (Hubl et al[30]). The region of the left middle frontal gyrus has been indicated to be related to central executive function (Frith et al[31]; D’Esposito et al[32]; Salmon et al[33]). In addition, Hoffman and Haxby[10]have reported that the intraparietal sulcus is related to eye-gaze perception in normal subjects, which is associated with spatial perception and spatially directed attention (Haxby et al[34]; Corbetta et al[35]; Nobre et al[36]; Corbetta[37]). Our AD patients may have undergone inappropriate emotional responses during the processing of the task used in this study, and, due to abnormalities in specific brain regions, they may have used strategies that differed from those of healthy subjects in order to perform the required task. Alternatively, the affective salience of the images may be relevant to the activation pattern seen in our autistic subjects. Insula activation is associated with processing of interoception, including somatosensory feedback associated with autonomic arousal (Davidson and Irwin[38]; Critchley et al[39]). One possible interpretation of the higher activation pattern of two TOM-related brain regions and the insula in the autistic subjects is that they may have been more affectively aroused by the images than the normal controls, such that feedback modulation of the gaze perception areas such as the superior temporal gyrus occurred. However, widely distributed impairment in the neural circuitry, including in the insula and the other three brain regions, may exist and bear the pathophysiology of autism. It is possible that such widely distributed abnormalities in brain function in AD patients may be reflective of a compensatory process. Our results suggest that processing the direction of another person’s eyes involves an element of perceiving mental states in others, and such a stimulus appears to be sufficient to disclose functional impairment in TOM-associated brain regions in high-functioning AD patients. Furthermore, our findings also indicate that high-functioning AD patients may, in a compensatory manner, recruit widely distributed brain regions that are not normally required to perform a socially-linked information processing task. |
||||
|
|
||||
Acknowledgment
|
||||
要旨
|
||||
References
|
||||
Others
|
||||
|